Les Arthropodes
Les Arthropodes du grec arthron « articulation » et podos « pied », aussi appelés « articulés » — sont un embranchement d'animaux protostomiens dont le plan d'organisation est caractérisé par un corps segmenté. Ils sont formés de métamères hétéronomes munis chacun d'une paire d'appendices articulés et recouvert d'une cuticule ou d'une carapace rigide, qui constitue leur exosquelette, dans la plupart des cas constitué de chitine. Leur mue permet, en changeant périodiquement leur squelette externe, de grandir en taille (mue de croissance) ou d'acquérir de nouveaux organes, voire de changer de forme (mue de métamorphose). Ils seraient apparus il y a 543 millions d'années.
Fiche récapitulative
- Métazoaires, triploblastiques, coelomates ;
- Symétrie bilatérale ;
- Protostomiens ;
- Métamérisés ;
- Squelette ectodermique chitineux, articulé ;
- Tube digestif complet formé de régions spécialisées ;
- Système circulatoire ouvert ;
- Système d’excrétion différencié ;
- Système nerveux formé de trois ganglions fusionnés et d’une chaîne nerveuse ventrale ;
- Marins, dulçaquicoles, ou terrestres ;
- Libres, sessiles, endo - ou ectoparasites ;
- Très grande diversité de modes de vie.

Présentation du groupe
Les Panarthropodes constituent le groupe qui comprend le plus grand nombre d’espèces décrites (environ 1 million), cela représente 85 % de la biodiversité connue. Ils dominent la terre depuis plus de 525millions d’années à la suite de l’explosion des formes de vie au Cambrien. Ils appartiennent aux Ecdyzozoaires. Leur tégument s’est épaissi par l’incorporation de protéines associées à un polysaccharide souple et résistant : la chitine. Cette modification fondamentale crée un exosquelette rigide, la cuticule, qui conditionne toute l’organisation interne de l’animal et l’oblige à passer par des mues (ecdysis) pour permettre sa croissance. Les mouvements seront rendus possibles par la division de la cuticule en plaques séparées. Arthropode signifie littéralement « pieds articulés ».
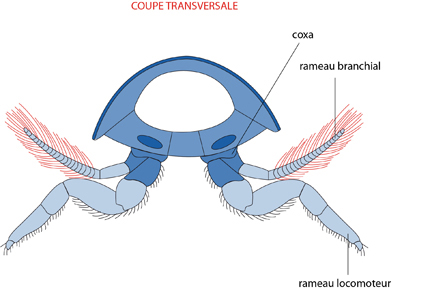
Les Panarthropodes
Les Panarthropodes sont fondamentalement des animaux segmentés.
Le modèle de base d’un segment peut-être déduit du schéma de la figure suivante.
Chaque segment porte une paire d’appendices biramés, c’est-à-dire composés d’un rameau branchial et d’un rameau locomoteur. Au cours de l’évolution, ces segments vont subir une spécialisation régionalisée selon deux processus : la perte de fonction de l’un des deux rameaux de l’appendice, et la fusion de certains segments. Ainsi, la tête est le résultat de la fusion d’une série de segments dont les appendices locomoteurs se sont transformés en appendices sensoriels ou en pièces buccales.
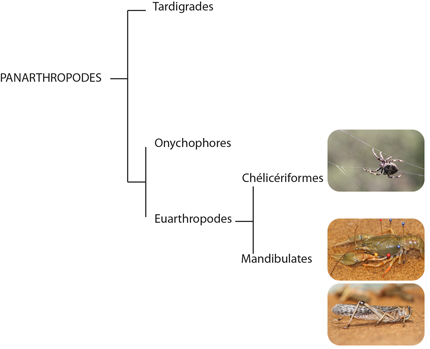
3 Clades
Les Panarthropodes se divisent en trois clades : Les Tardigrades, les Onychophores et les Euarthropodes. Les Onychophores et les Euarthropodes partagent un système circulatoire ouvert avec un cœur segmenté présentant des ostia, une hémocyanine semblable, une musculature des membres segmentée, et des néphridies. Les Tardigrades semblent plus éloignés sur le plan phylétique. Nous nous focaliserons principalement sur les Euarthropodes eux-mêmes divisés en Chélicériformes et en Mandibulates. Les organismes modèles choisis sont l’écrevisse (Pancrustacé Malacosracé), le criquet pèlerin (Pancrustacé Hexapode) et l’Epeire diadème (Chélicérate, Arachnide).

2. Exemple-type 1 : L’écrevisse (Astacus astacus), Malacostracé - 2.1. Examen externe
Astacus, l’Ecrevisse est représentés sur la photo suivante. Astacus vit dans les rivières bien oxygénées et y joue un rôle de prédateur. Traditionnellement, il existait deux méthodes pour la capturer. La première est un piège conçu pour cette pêche ; la seconde est beaucoup plus rustique : on utilise une vulgaire baguette de bois que l'on fend à son extrémité. Dans cette fente, on coince l'appât : un morceau de viande ou de poisson, et on le présente silencieusement devant un trou de la berge susceptible de servir de refuge à une Ecrevisse. Si c'est le cas, le crustacé ne manque pas de sortir de son trou, et commence à s'intéresser au repas qui lui est offert. Après quelques instants, lorsque l'Ecrevisse est occupée à manger, on retire lentement la baguette. Voyant son repas lui échapper, elle s'y agrippe avec ses pinces et il ne reste plus au pêcheur qu'à l'amener sur la berge.

Astacus astacus
L'Ecrevisse autochtone, Astacus astacus, appelée plus simplement écrevisse à pattes rouges, a pratiquement disparu de nos rivières. A la fin du siècle dernier, elle a été victime d'un petit champignon parasite, que l'on connaît sous le nom de « peste de l'écrevisse ». A cette épidémie, il faut ajouter la pollution des eaux, mais également les curages et les rectifications des berges dont sont victimes nos rivières ! Dans ces conditions, comment trouve-t-on des écrevisses dans les poissonneries? Simplement grâce à l'astaciculture: les pisciculteurs produisent ces crustacés de façon industrielle. Il s'agit d'espèces étrangères: écrevisses de Russie, ou américaines. Echappées des astacicultures, ces espèces allochtones ont envahi les cours d'eau et deviennent invasives, éliminant les derniers représentants de l’espèce locale qui pouvaient y subsister.
Le corps de l'Ecrevisse est protégé par une cuticule imprégnée de sels calcaires. Cette cuticule minéralisée forme une armure solide, articulée, qui laisse la liberté de mouvements aux appendices, articulés et caparaçonnés eux aussi.
Elle est divisée en anneaux inégalement distincts disposés longitudinalement. Dans la partie antérieure du corps, plusieurs anneaux sont réunis en un tronçon rigide formant la carapace qui protège le céphalothorax. Les anneaux postérieurs sont distincts et mobiles formant l'abdomen souvent replié sous le corps.
L'Ecrevisse présente une série de paires d'appendices, plus ou moins allongés, et apparemment bien compliqués. Fondamentalement pourtant, tous ces appendices sont semblables. Les différences observées sont le résultat d'adaptations secondaires à des fonctions diverses.
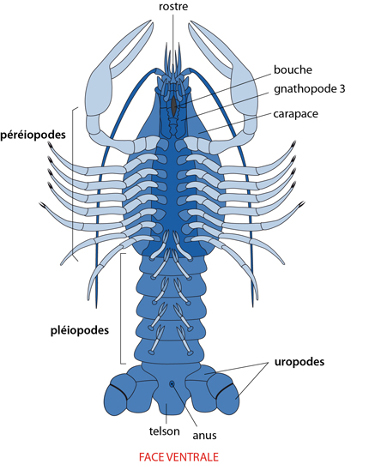
Le Telson
En général, il y a 20 paires d’appendices, plus un petit rostre à l'avant (formé par l'acron) et une plaque impaire au bout de l'abdomen formant le telson. De part et d'autre du telson, la dernière paire d'appendices abdominaux est transformée en uropodes servant à la nage. Les 5 paires qui précèdent s'appellent pléiopodes, sont peu développées et ont une fonction sexuelle. Plus en avant, et articulées au thorax, les 8 paires d'appendices se dénomment péréiopodes. Parmi ceux-ci, 5 paires de grandes pattes sont vouées à la locomotion. En se rapprochant de la partie antérieure du corps, la paire péréiopode restante a une fonction préhensile: la pince qui est diantrement puissante. En avant encore, 3 paires de pattes-mâchoires ou gnathopodes, plus petites, sont affectées à la mastication. Sur le schéma n'est visible que la dernière des gnathopodes, les « gnathopodes 3 ». Ils recouvrent les deux autres paires.
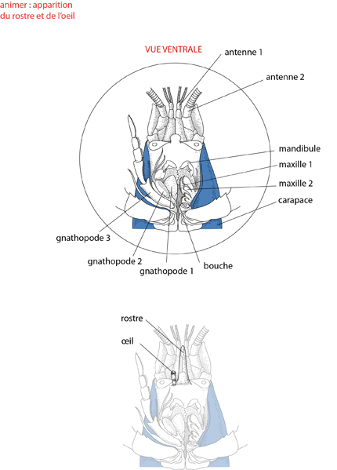
Les pièces buccales
Détaillons les pièces buccales. On retrouve les gnathopodes, les paires de maxilles, et la paire de mandibules qui les précèdent. Situés en avant de la cavité buccale, on trouve les appendices sensoriels: 2 paires d'antennes (une longue et une courte), et une paire d'yeux portés par les pédoncules situés de part et d'autre du rostre, dans une échancrure de la carapace. L'œil, plus dorsal que les antennes, a été dégagé à gauche sur le dessin. Ces appendices apparaissent effectivement fort variés. Bâtis sur le même plan fondamental? Ce n'est guère évident à première vue; pourtant, si! La structure de base dont ils dérivent est représentée de façon satisfaisante par la troisième paire de gnathopodes que vous découvrez dans le dessin suivant.
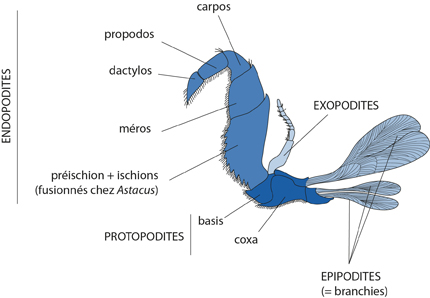
Appendice biramé
C'est un appendice biramé. Une partie basale, le protopodite porte 2 rames terminales: l'endopodite ou rame interne, et l'exopodite ou rame externe. Le protopodite porte des branchies et correspondent au rameau branchial. Au cours de l'évolution des Crustacés, des expansions lamellaires plus ou moins complexes ont garni l'un ou l'autre segment du protopodite, et ont servi à des fonctions diverses: aider à la mastication, à la nage, au transport des œufs. A ces expansions, on a donné le nom général d'épipodites. Le basis du protopodite porte les deux rames. La rame externe, ou exopodite comporte un segment basal et un distal constitué de nombreux articles. Ce plan de base, illustré avec les troisièmes gnathopodes d'Astacus, varie presqu'à l'infini chez les Crustacés. Il est déjà très modifié sur les autres appendices de l'Ecrevisse, en fonction du rôle qu'ils ont à remplir.
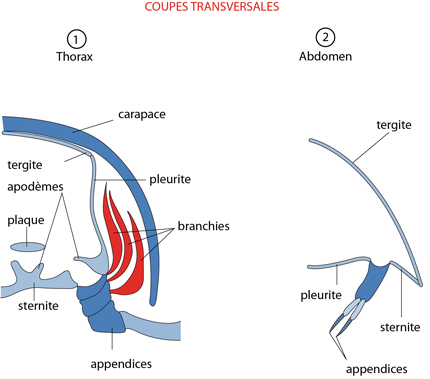
2.2. Examen interne - 2.2.1. Tégument
Revenons à la cuticule protégeant le corps, pour fixer un peu plus de ce vocabulaire complexe. Chaque segment de l'Ecrevisse est muni de 4 plaques : une tergite dorsale, une sternite ventrale, et 2 pleurites latérales, sur lesquelles sont insérées les appendices. La carapace déborde latéralement largement du corps formant une chambre branchiale ouverte vers le bas. Ainsi cette carapace ménage un espace entre sa face interne et le pleurite, dans lequel sont déployées les branchies délicates. Les mouvements sont possibles puisque cette cuticule est disposée en plaques séparées. Les plaques sont régulièrement disposées à chaque segment, comme dans la région abdominale, et les plaques d'un segment sont jointes à celles du segment suivant par une membrane articulaire, c'est-à-dire une zone où la cuticule reste mince et flexible. Cette disposition régulière disparaît fréquemment au cours de l’évolution, via des fusions ou de subdivisions secondaires. Comme l'exosquelette du corps, celui des appendices est divisé en sections tubulaires réunies les unes aux autres par des membranes articulaires; il y a donc une articulation à chaque jonction. Ces dispositifs permettent aux sections des appendices, comme aux segments du corps, de se mouvoir l'un par rapport à l'autre.
En plus de l'exosquelette, un « endosquelette » s'est parfois développé. Il s'agit d'invaginations de la cuticule, produisant des projections internes ou apodèmes. En outre, des calcifications de tissus internes forment des plaques libres. Si vous avez mangé des Ecrevisses ou autres Crustacés, vous avez sans doute remarqué, en arrachant un appendice, que vous entraîniez une fine lame plate, assez longue, qui doit bien se loger quelque part dans la profondeur des muscles du thorax ou de l'abdomen. La présence de l'endosquelette, d'apodèmes et de plaques permet l'insertion des muscles.
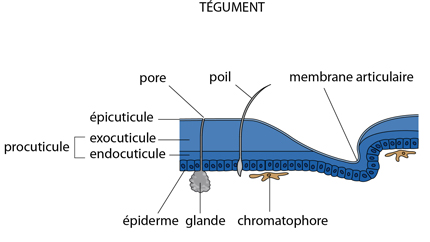
L’exosquelette
L’exosquelette (à part les plaques internes) est sécrété par une couche de cellules épidermiques. Il se compose d'une épicuticule externe et d'une procuticule beaucoup plus épaisse. L'épicuticule est constituée de protéines. La procuticule se divise en exocuticule externe et endocuticule interne. Ces couches contiennent des protéines et de la chitine, associées pour former des glycoprotéines complexes. L'exocuticule est absente des membranes articulaires, ainsi que le long de lignes de moindre résistance, où l'exosquelette craquera lors de la mue. Chez les Crustacés comme l'Ecrevisse, la procuticule est imprégnée de phosphate et de carbonate de calcium.
Aux endroits amincis, la cuticule de l'Ecrevisse est perméable aux gaz et à l'eau. Elle est aussi percée de fins canalicules par lesquels transitent les sécrétions des cellules glandulaires épidermiques.
Les différentes parties de la cuticule sont produites par les cellules épidermiques et sont donc des dérivés ectodermiques.

Le tégument
Les couleurs du tégument résultent du dépôt de pigments dans la cuticule, mais aussi de la présence de cellules spéciales, les chromatophores, situées sous l'épiderme. Les chromatophores sont branchus, étoilés; les grains de pigments peuvent se disperser dans les prolongements, ou au contraire se concentrer au centre de la cellule; dans cet état, la coloration sera moins visible. Sur l'animal vivant, ce jeu de pigmentation permet le camouflage, l'éclaircissement et l'assombrissement selon « l'humeur » du Crustacé.
La photo suivante suggère une particularité de ces pigments. Vous avez aussi remarqué, en mangeant votre Homard, votre Crabe ou votre Langouste, que sa couleur avait changé pendant la cuisson: ces Crustacés deviennent rouges parce que leur pigment bleu est dû à l'association du caroténoïde avec une protéine et que cette association est détruite à la chaleur.

Activité locomotrice
L'exosquelette combine des avantages pour l'activité locomotrice et pour la protection, mais en revanche, il pose des problèmes à l'animal en terme de croissance. La solution, c'est l'évacuation périodique du vieil exosquelette trop étroit, à la mue appelée aussi ecdysis. La photo suivante vous montre un exosquelette abandonné, on l’appelle exuvie. Avant d'ôter son vieil habit, l'Ecrevisse se prépare. Les cellules épidermiques se détachent de l'exosquelette et sécrètent une nouvelle épicuticule. Puis elles sécrètent des enzymes qui traversent la nouvelle épicuticule et vont éroder la vieille endocuticule, en respectant les attaches musculaires et nerveuses, si bien que l'animal peut continuer à se mouvoir. Après la digestion de l'endocuticule, l'épiderme secrète une nouvelle procuticule. Le vieux squelette craque maintenant le long de certaines lignes prédéterminées, et l'Ecrevisse s'en extirpe. Le nouvel exosquelette est encore mou, et tout fripé. L'Ecrevisse s'y met à l'aise, et le minéralise en utilisant une réserve de sels de calcium qu'elle s'était constituée dans l'estomac. A. astacus mue une vingtaine de fois pendant les 4 premières années de sa vie, avant d'atteindre la maturité sexuelle. Les stades entre les mues sont dénommés instars, et ils deviennent de plus en plus longs au fur et à mesure que vieillit l'animal.
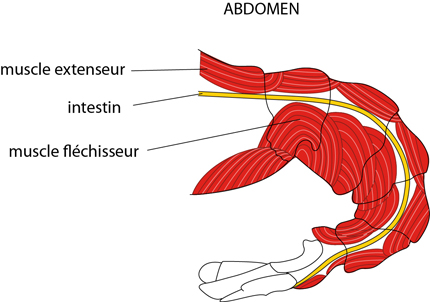
2.2.2. Système locomoteur
Les mouvements de l'Ecrevisse consistent en flexions entre les plaques de l'exoquelette. Les muscles s'attachent à la surface interne de la procuticule par des fibrilles. Les flexions et les extensions entre les plaques proviennent de la contraction de ces muscles.
Chez l'Ecrevisse armée d'un céphalothorax rigide, les mouvements se réduisent à ceux de l'abdomen et des appendices. Prenons l'abdomen, par exemple: le groupe de muscles extenseur est dorsal, et le groupe de muscles fléchisseurs est ventral; c'est lui qui fait se replier violemment la partie postérieure de l'abdomen sous le corps lorsque l'Ecrevisse veut s’échapper, à reculons.

2.2.2. Système locomoteur
Cette action combinée du squelette et de la musculature est fort comparable à celle des Vertébrés; la différence principale c'est que les muscles des Arthropodes sont attachés à la surface interne de l'exosquelette tandis que les muscles des Vertébrés sont attachés à la surface externe d'un endosquelette. Pour certains mouvements, des appendices notamment, l'exosquelette devient d'un emploi difficile. Les apodèmes et les plaques rigides internes facilitent alors des insertions musculaires plus efficaces.
L'entièreté de la musculature de l'Ecrevisse, comme celle des autres Arthropodes, est constituée de cellules musculaires striées, même autour des organes internes. On ne trouve aucune cellule musculaire lisse.
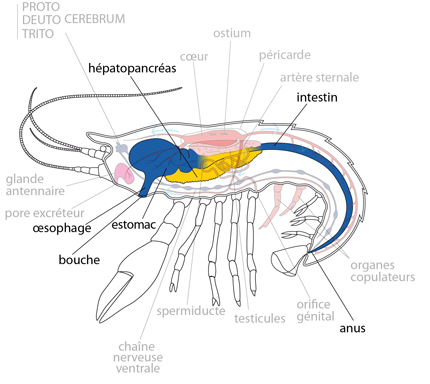
2.2.3. Système digestif
Examinons le processus de nutrition au travers du schéma suivant. Par ses pinces, l'Ecrevisse a saisi sa pitance. C'est un carnivore très peu attentif à la fraîcheur de sa nourriture. Elle porte un morceau à sa bouche, fendue longitudinalement et entourée d'un appareillage masticateur complexe: la paire de mandibules, les 2 paires de maxilles et, comme si c'était encore insuffisant, et les 3 paires de gnathopodes. L'œsophage, très court et presque vertical, est la première portion du tube digestif.
Le tube digestif présente des régions stomodéale (antérieure) et proctodéale (postérieure) très longues, colorées en bleu sur le dessin. Ces parties sont pourvues d'une cuticule et sont donc d’origine ectodermique. En cela, le tube digestif des arthropodes diffère de celui de la plupart des autres animaux. Seule la partie moyenne, ou mésentéron possède un épithélium d'origine endodermique, ici en jaune.
Le stomodeum
Le stomodeum joue un rôle dans l'ingestion, la trituration et le stockage de la nourriture. L'œsophage, non visible sur la photo, débouche dans l'estomac, très volumineux. Un étranglement divise l'estomac en deux parties; dans la partie antérieure, on trouve deux masses calcaires plus ou moins saillantes. Elles constituent une provision de sels calcaires, que l'animal utilise après sa mue, pour former sa nouvelle carapace.

L'estomac
L'estomac porte un ensemble de pièces cuticulaires résistantes, des dents qui forment le moulin gastrique, ici en jaune sur la photo. Elles broient une fois encore les aliments, poursuivant ainsi la digestion extracellulaire mécanique.
Le mésentéron
Le mésentéron est la partie du système digestif où se poursuit la digestion enzymatique, et l'absorption. Deux volumineuses glandes digestives jaunes-citron, forment « l’hépato-pancréas » par comparaison assez judicieuse avec les organes de même nom chez les Vertébrés. Ces glandes sont des évaginations très ramifiées de l'épithélium du mésentéron. L'hépato-pancréas est le site de production des enzymes digestifs. Ces enzymes sont déversées dans l'estomac par deux canaux, où s'opère la digestion chimique extra-cellulaire des aliments. Les nutriments sont ensuite absorbés par les cellules du mésentéron, c'est-à-dire une très courte portion rectiligne qui fait suite à l'estomac, et les cellules de l'hépato-pancréas lui-même. Celles-ci sont capables de stocker du glycogène, des graisses et des sels de calcium.

Le proctodeum
Le proctodeum est le siège de réabsorption d'eau et d'ions, ainsi que de la formation des matières fécales. Chez l'Ecrevisse, c'est un long intestin rectiligne parcourant toute la longueur de l'abdomen, pour aboutir à l'anus, situé juste devant le telson.
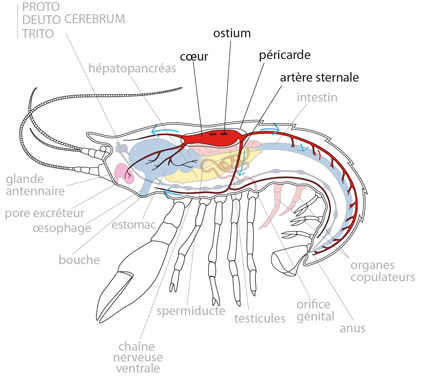
2.2.4. Système circulatoire
Les nutriments absorbés par le mésentéron passent dans le sang. Nous allons examiner maintenant le système circulatoire.
Chez les Arthropodes, les métamères ne sont pas divisés par des septa. Il en résulte une large cavité centrale appelée hémocœle où baignent les organes. Il s’agit d’espaces inter-tissulaires remplis de liquide, que l’on appelle hémolymphe, puisqu’il est constitué à la fois du sang (hémo) et de la lymphe qui baigne les espaces intercellulaires. Le cœlome métamérique analogue à celui des Annélides n’est représenté que par les cavités des gonades, et, chez l'Ecrevisse, par une portion des organes excréteurs. Cette réduction est liée au mode de locomotion: le squelette hydrostatique a fait place à un solide squelette externe.
Le système vasculaire sanguin est ouvert. Le cœur et le vaisseau dorsal assurent la propulsion du sang. Le cœur est une chambre musculeuse, percée de paires d'orifices latéraux, dénommés ostioles. Les ostioles permettent à l'hémolymphe de pénétrer dans le cœur à partir du grand sinus qui l'entoure, et qui porte le nom de cavité péricardique.
L'hémolymphe
L'hémolymphe est expulsée du cœur par sa contraction. Grâce à un système de valves anti-retour, l'hémolymphe est propulsée dans des artères; vers l'avant dans les artères dorsales antérieures, et vers l'arrière dans l'artère dorsale postérieure. Elle parvient aussi dans l'artère ventrale via l'artère sternale. Dans certaines régions du corps, les artères se prolongent par des vaisseaux, par exemple dans les appendices thoraciques et les branchies. Les artères peuvent aussi se prolonger par des capillaires, par exemple autour du mésentéron, où l'hémolymphe se charge de matières nutritives, ou autour du proctodeum. De toute manière, l'hémolymphe finit par se répandre dans les sinus qui baignent les organes. Elle les nourrit et les oxygène. Elle revient ensuite au cœur par les ostia.

2.2.5. Système respiratoire
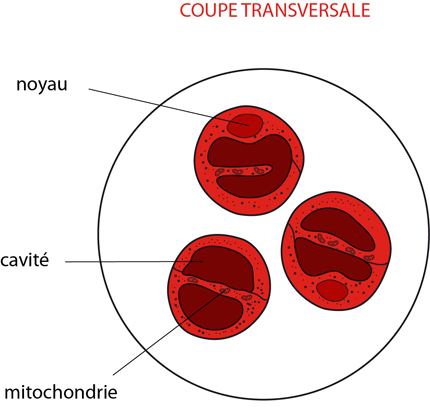
Le système respiratoire est intimement lié au système circulatoire comme le montre cette coupe transversale faite au niveau du cœur. L'hémolymphe en provenance des branchies rejoint la cavité péricardique, et rentre dans le cœur par les ostioles. Les flèches indiquent le sens de la circulation.
Les branchies sont attachées aux appendices, et protégées par une expansion de la cuticule. Donc, de part et d'autre du corps, se délimite une cavité branchiale ouverte vers le bas, et dans laquelle l'eau circule constamment. Les branchies plumeuses sont constituées d'une touffe de minces filaments portés par une sorte d'axe. La paroi des filaments est épidermique et revêtue d'une mince cuticule perméable.
L'hémolymphe en provenance des sinus circule dans ces filaments. L'hémolymphe contient des pigments respiratoires en solution. Chez l'Ecrevisse, ce pigment est l'hémocyanine, gros polymère protéique doté de groupes hémiques comme l'hémoglobine, mais où le fer est remplacé par du cuivre. L'hémocyanine fixe l'oxygène, comme l'hémoglobine. En passant par les filaments branchiaux, l'hémolymphe se chargera donc d'oxygène en provenance de l'eau ambiante. D'autre part, l'hémolymphe se déchargera du CO2 qu'elle véhicule sous forme d'ions dicarbonates.
Question
Un petit problème simple de morphologie fonctionnelle : si l'on opère une coupe transversale dans les filaments branchiaux d'A. astacus, on constate, dans la plupart des sections, la présence d'une cloison disposée comme le montre ce schéma. Pourquoi ?

2.2.6. Système excréteur
Osmorégulation et excrétion : chez la plupart des animaux, ces deux fonctions sont couplées. Les branchies, visibles ici sur la photo, ont d’autres fonctions outre la respiration: elles participent activement à l'excrétion d'un autre déchet du catabolisme, l'ammoniaque, qui est effectivement le principal détritus azoté chez les Crustacés. Les déchets azotés et le CO2 ne sont pas les seuls produits du catabolisme à excréter. Des molécules ou des particules plus volumineuses peuvent s'accumuler dans des cellules spéciales, appelées néphrocytes, circulant dans l'hémolymphe, puis se rassemblant à certains endroits du corps, notamment à la base des branchies et des appendices, qui les excrètent.
Les branchies interviennent aussi dans l'osmorégulation. Chez l'Ecrevisse, qui vit en eau douce, elles absorbent activement le peu de sels que l'animal peut pomper dans son milieu ambiant.
Pouvez-vous expliciter quelle serait le rôle des branchies dans la régulation osmotique chez un Crustacé marin ?
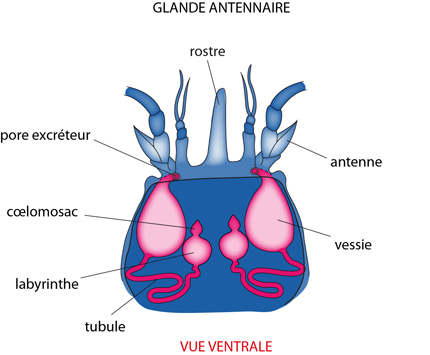
Les glandes antennaires
Un autre organe est voué à l'osmorégulation et à l’excrétion chez l'Ecrevisse : les glandes antennaires. Ces deux glandes sont localisées dans la partie antérieure du céphalothorax; elles filtrent des déchets de l'hémolymphe circulant dans les sinus proches, puis évacuent les déchets par deux orifices excréteurs, situés à la base des antennes (d'où le nom: glandes antennaires). Si ces glandes étaient seules à intervenir dans l'excrétion, au vu de leur situation et des particularités du système circulatoire, le résultat serait inefficace. Cependant, elles travaillent de concert avec les branchies et les néphrocytes.
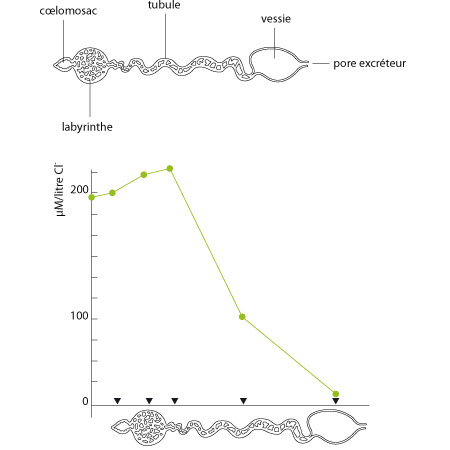
Le coelomosac
Chaque glande antennaire comprend au bout aveugle un petit sac dérivé du cœlome, le «cœlomosac». Un dispositif d'ultrafiltration somme toute assez comparable à l'ensemble lame basale-podocytes du rein des Mammifères, constitue sa paroi. (Si vous avez oublié de quoi il s'agit, allez revoir votre cours de première candi!). L'urine filtrée se retrouve dans la cavité du cœlomosac, et sera modifiée en passant dans la portion suivante de la glande antennaire: le labyrinthe, puis le tubule. L'urine définitive est stockée dans la vessie et évacuée périodiquement par le pore excréteur.
2.2.7. Système nerveux
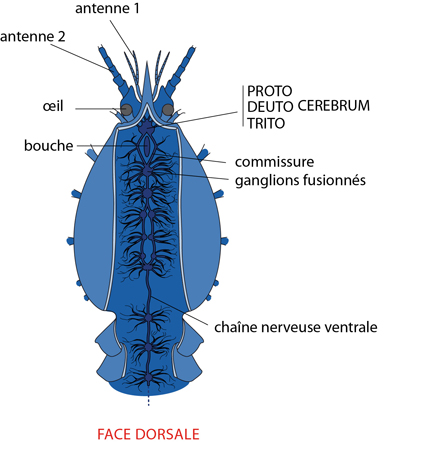
Les activités de l'Ecrevisse sont coordonnées par le système nerveux et par le système endocrinien. Comme chez les autres Arthropodes, la céphalisation est importante: l'augmentation de taille du cerveau est corrélée à des organes des sens bien développés, tels les yeux et les antennes, et le comportement est relativement complexe.
Le cerveau est formé de trois masses fusionnées, chacune gardant des traces d'une origine paire: un protocerebrum antérieur, un deuterocerebrum médian et un tritocerebrum postérieur. Comme la masse cérébrale est repliée vers l'arrière, cette séquence semble inversée sur le dessin. Nous détaillerons le système nerveux céphalique des arthropodes avec l’exemple du criquet
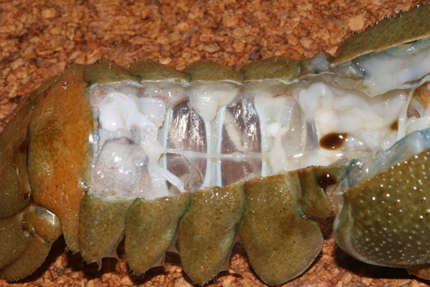
Chaine nerveuse d’écrevisse
Voici une chaine nerveuse d’écrevisse, en place. Vous pouvez déjà y repérer les ganglions.
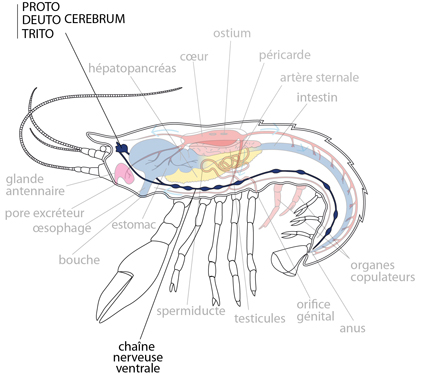
Chaine nerveuse d’écrevisse
Une particularité : la chaîne nerveuse est parcourue d'axones géants, comme chez les Annélides ; ils fonctionnent un peu comme ces derniers : une décharge stimule violemment la contraction de la musculature abdominale ventrale en cas de danger, et l'Ecrevisse replie son abdomen et s'enfuit à reculons.
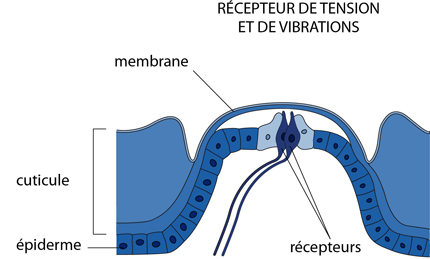
L'exosquelette
L'exosquelette est très avantageux pour la protection et la locomotion, mais par contre, il limite la perception sensorielle en formant une barrière à la détection des stimuli extérieurs. Les Arthropodes ont résolu ce problème de manière souvent très originale et très performante. Pour ce faire, les récepteurs sensoriels sont associés à des modifications locales de l'exosquelette chitineux. Des interruptions de l'exosquelette sont couvertes d'une fine membrane, sous laquelle sont connectés des récepteurs sensoriels. Ces organes détectent les vibrations de la membrane, ou d'autres forces qui changent la tension du squelette.
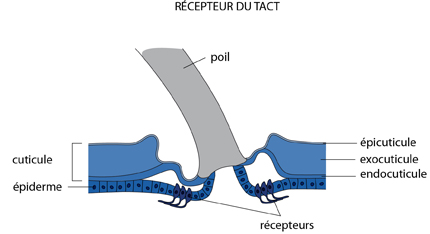
Le récepteur
Un premier type de récepteur, important et très commun, présenté dans le schéma suivant, est connecté aux poils ou aux soies qui garnissent l'exosquelette: lorsque le poil bouge, des terminaisons réceptrices localisées à sa base sont stimulées ; ainsi se résout le problème du tact.
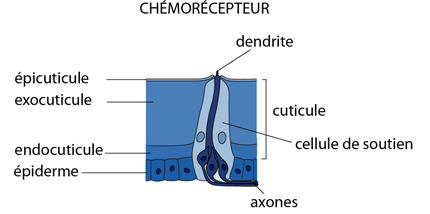
Les chémorécepteurs
Des chémorécepteurs sont logés au fond de canaux, de fentes, qui traversent l'exosquelette, ou encore à la surface de petits poils spéciaux comme dans le schéma.
Tous ces types de récepteurs sensoriels peuvent être dispersés sur la surface du corps, ou alors rassemblés en des endroits privilégiés: les articulations, les pattes, les palpes, les antennes.
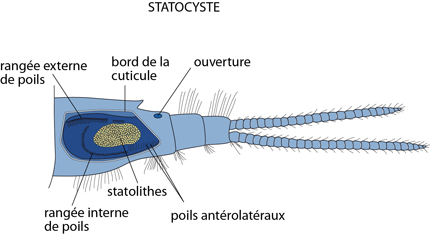
Les statocystes
L'Ecrevisse est renseignée sur sa position dans l'espace lors d’un déplacement par une paire de statocystes, localisés dans l'article de base des antennules. Les statocystes sont de simples invaginations sphériques de l'exosquelette, dont le creux contient des grains de sable agglomérés par les sécrétions de la paroi du sac. Cette paroi est tapissée de petits poils sensoriels. Selon la position de l'animal, les grains de sable se déposeront sur telle ou telle partie de la surface interne du statocyste et stimuleront donc les poils correspondants. Puisque le statocyste est une invagination de l'exosquelette, la cuticule est évacuée à chaque mue avec le contenu. Chaque fois, l'Ecrevisse doit donc y réintroduire des grains de sable.
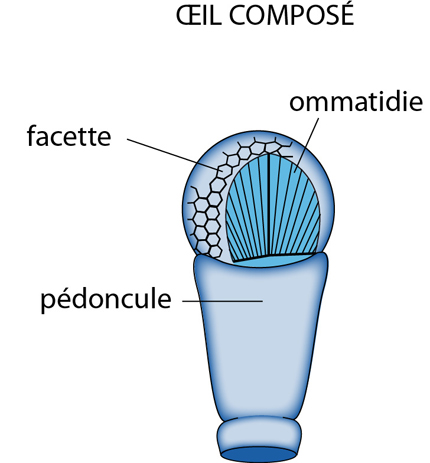
L'oeil
Mais l'organe des sens le plus remarquable, c'est l’œil ou photorécepteur.

Les yeux
L'Ecrevisse possède 2 yeux portés par un pédoncule, et logés dans une échancrure de la carapace, de part et d'autre du rostre. L'œil est composé d'unités cylindriques possédant chacune tous les éléments nécessaires à une bonne photoréception.
Chaque unité cylindrique, dénommée ommatidie, est couverte extérieurement par une cornée transparente dérivée de la cuticule. La cornée fonctionne comme une lentille. La surface externe de la cornée, la facette, présente un contour hexagonal.
Sous la cornée, l'ommatidie comporte un cône cristallinien, qui fonctionne comme une seconde lentille. Ce cône est produit par 4 cellules.
Les éléments photorécepteurs proprement dits sont localisés juste en-dessous du cône. Ils forment la rétinule. Le centre de la rétinule est occupé par un cylindre transparent, le rhabdome, autour duquel se disposent les cellules photoréceptrices allongées. On dénombre 8 cellules photoréceptrices. La surface interne de chacune est garnie de microvillosités orientées perpendiculairement à l'axe du rhabdome. La membrane plasmique tapissant les microvillosités renferme les pigments photo-sensibles. Les 8 ensembles de microvillosités se font face et constituent le rhabdome. Les cellules photoréceptrices sont en fait des neurones très modifiés: leurs dendrites correspondent à la zone porteuse de microvillosités, et leurs axones se dirigent vers les ganglions optiques localisés dans les pédoncules oculaires; l'information y est analysée, puis transférée vers les centres optiques du protocerebrum.
Dans la périphérie de l'ommatidie, on trouve encore des cellules pigmentaires, dont le cytoplasme contient des grains noirs. Elles forment un manchon opaque, un écran, si bien que la lumière pénétrant dans une ommatidie stimulera seulement le rhabdome de ladite ommatidie, et pas les voisins. Cependant, ce système est réglable. Si la lumière est faible, les pigments peuvent migrer le long du cytoplasme des cellules pigmentaires, et laisser passer la lumière d'une ommatidie à l'autre.
L'image totale élaborée par un œil composé résulte de la stimulation d'ommatidies individuelles. En première approximation, on pourrait comparer cette image à celle d'un téléviseur, sur l'écran duquel la juxtaposition des points lumineux forme l'image. En fait, la rétine des Vertébrés fonctionne de la même manière: l'image résulte de la stimulation des cônes et bâtonnets individuels, mais son grain est plus fin.
L'image fournie par l'œil composé est moins bien définie, mais ce type d'œil apporte deux avantages différents: d'abord sa construction se prête bien à la vision dans un champ visuel très large: la surface peut couvrir un angle de 180° ou plus. En second lieu, l'œil composé détecte très facilement le mouvement.
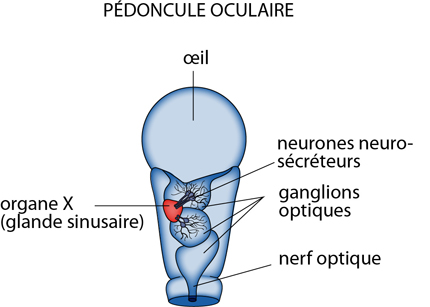
2.2.9 Système endocrinien
Chez l'Ecrevisse, et les Crustacés en général, on trouvera des neurosécrétions (c-à-d des hormones sécrétées par des cellules nerveuses, comme celles sécrétées par les neurones hypothalamiques des Vertébrés), et des hormones produites notamment par 3 tissus endocrines: l'organe Y, la glande androgène et l'ovaire. Des groupes de neurones neurosécréteurs sont localisés à divers endroits, dans les ganglions optiques des pédoncules oculaires. Les produits synthétisés par les corps cellulaires migrent vers la glande sinusaire ou organe X, située contre les ganglions optiques, et qui sert de lieu de stockage. De là, les neurosécrétions sont libérées dans l'hémolymphe. Elles vont contrôler les sécrétions hormonales des autres tissus endocrines, notamment lors de la mue.

Les hormones
Les neurosécrétions et les hormones de l'Ecrevisse règlent des fonctions très diverses, y compris des aspects variés du métabolisme général. Nous nous contenterons d'illustrer le contrôle de deux phénomènes majeurs: la mue et la reproduction. On sait déjà que l'Ecrevisse doit muer périodiquement au cours de sa croissance. Tant que la mue n'est pas nécessaire, les neurones neurosécréteurs inhibent les sécrétions hormonales de deux organes Y. Lorsque cesse l'inhibition, les organes Y, localisés à gauche et à droite dans le céphalothorax, produisent une hormone, l'ecdystérone. L'ecdystérone stimule alors les cellules concernées par la mue: les cellules de l'hépatopancréas qui accumulent des réserves, et les cellules épidermiques qui recyclent notamment le calcium de l’ancienne cuticule.
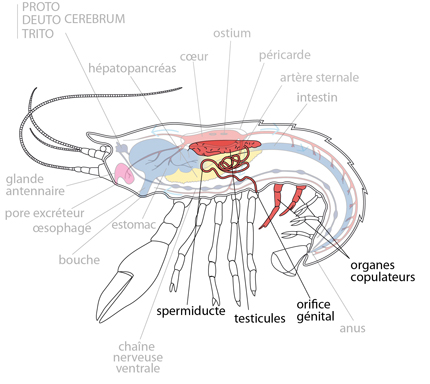
2.2.10. Système reproducteur
L'Ecrevisse est dioïque. Les gonades sont placées dorsalement, en-dessous du cœur, et elles sont condensées en une seule masse. Chez le mâle, les testicules déversent les spermatozoïdes par deux spermiductes très tortueux qui aboutissent aux orifices génitaux, situés sur l'article basal des pattes de la cinquième paire thoracique. Grâce aux premières paires de pléiopodes abdominaux modifiées en organe copulateur, le mâle dépose ses spermatozoïdes englués dans les sécrétions du spermiducte, près des orifices génitaux de la femelle.
Les ovules
Les ovules quittent les ovaires par deux oviductes et parviennent aux orifices localisés sur l'article basal de la troisième paire de pattes thoraciques. Notons que tous les pléiopodes abdominaux sont semblables, sauf la première paire, qui est atrophiée et permet ainsi le jeu des organes copulateurs mâles. Les ovules sont fécondés dès qu'ils surgissent de l'orifice génital femelle. Ils sont entourés d'une coque résistante et gluante et attachés aux pléiopodes de la femelle. Là, les œufs se développent à l'aise, bien protégés par leur mère et bien oxygénés avec les battements continus des pléiopodes. À l'éclosion, une petite écrevisse miniature sort de l'œuf.
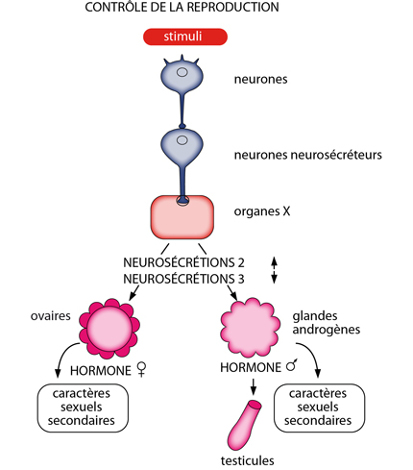
Les gonades
Le développement des gonades, des caractères sexuels secondaires (en particulier les différences des appendices entre mâles et femelles), ainsi que la période de reproduction sont contrôlés par les hormones.
Les neurosécrétions (toujours via l'organe X) inhibent le développement des ovocytes en-dehors des périodes de reproduction. Lorsque la saison est favorable, cette neurosécrétion décline. Une autre neurosécrétion, stimulante cette fois, apparaît et les œufs se développent. Pareillement, chez le mâle, les neurosécrétions contrôlent le fonctionnement des glandes androgènes situées au bout des spermiductes. Les hormones de ces glandes stimulent les testicules et l'apparition des caractères sexuels mâles. A titre de curiosité: bien sûr, le sexe est déterminé génétiquement mais si l'on extirpe les glandes androgènes, les testicules se transforment en ovaire, et les caractères mâles disparaissent. Inversement, si l'on greffe des glandes androgènes chez des femelles, celles-ci se transforment en mâles fonctionnels. Donc les hormones de ces glandes servent d'intermédiaire pour la détermination du sexe, entre le génome et le phénotype.

3. Exemple type 2 : Le criquet Locusta migratoria (Hexapode)
Notre second exemple-type de l'Embranchement des Arthropodes, nous le prenons dans la Classe des Insectes. Les Insectes sont aussi appelés « hexapodes » parce qu’ils possèdent 6 pattes au stade adulte. Voici le Criquet migrateur appelé Locusta migratoria, ou encore Locuste. Ce Criquet migrateur marche, saute, vole. Il existe sous une forme solitaire et sous une forme grégaire qui peut effectuer des déplacements sur des distances considérables de plusieurs centaines, voire milliers de kilomètres. Le passage d’une forme à l’autre est lié aux conditions climatiques. Après de fortes pluies en région aride, la végétation se développe brutalement. Les criquets disposent alors d’une abondance de nourriture et se multiplient, les densités de la population augmentent et ils se transforment en forme grégaire. Avec leur multiplication et la réapparition de la sécheresse, la végétation diminue à nouveau et les criquets se concentrent alors dans les seules zones restées vertes. Quand la densité de la population diminue la forme grégaire peut se retransformer en forme solitaire.

Les criquets
Sous l’influence des contacts entre individus, les criquets subissent des transformations hormonales et la forme grégaire apparaît après deux ou trois générations, capable de voler en essaim sur de longues distances et de causer en quelques heures des dégâts irréparables aux cultures. La photo montre un rassemblement de Criquets sur un arbuste; ils l'ont complètement défeuillé.
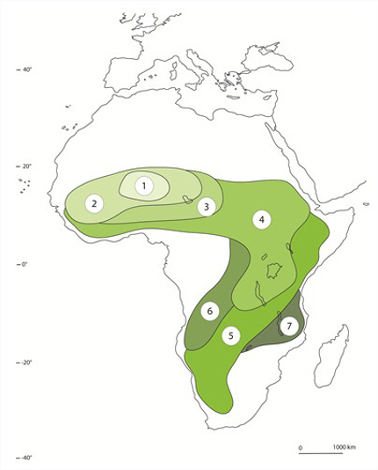
Les criquets
En 1928, des Criquets partis du Niger, signalé par le chiffre 1, ont envahi progressivement le Sud du Sahara, qu'ils ont dévasté, puis l'Afrique centrale. Ce genre de migration massive se reproduit périodiquement, même de nos jours. La dernière invasion date des années 2003 à 2005 et a nécessité le déploiement de moyens considérables et la pulvérisation de milliers de litres d’insecticides pour un coût estimé à 500 millions de dollars. Les bandes de Criquets migrateurs comptent des milliards d'individus, qui sont un véritable fléau pour l'agriculture. Un essaim mange l’équivalent de la nourriture de 2500 personnes en une journée. On parle de leurs ravages depuis la plus haute antiquité. Ils furent sans doute l'une des dix plaies d'Egypte décrite dans la Bible. L. migratoria est une espèce africaine, mais d'autres très proches dévastent l'Europe, l'Orient, l'Amérique. Actuellement, les organisations internationales se penchent sur ce problème; des chercheurs étudient les traits d’histoire de vie des Criquets, leurs lieux de rassemblement et leurs aires de déplacement, dans le but de contrôler leurs formidables accroissements de population et de prévenir les désastres qu'ils occasionnent.

Le criquet
Ici, un Criquet en phase solitaire. Sa livrée est de couleur claire et assez terne, son mode de vie est sédentaire, et il ne représente pas de danger pour l’agriculture. Sa fécondité est faible et il se déplace peu.

Le criquet
La phase grégaire se traduit par des modifications du comportement et de l'anatomie. Ainsi cette photo montre à quel point la livrée a changé ! Elle est plus sombre et les couleurs plus variées. Leur fécondité est plus élevée. Comme les adultes, les larves grégaires montrent des couleurs plus variées. En outre, leur développement est accéléré: elles parviennent plus vite à l'âge adulte et se reproduisent aussitôt.
Pourtant, il s’agit bien de la même espèce : les individus de la forme grégaire et de la forme solitaire peuvent se reproduire entre eux et former des descendants viables. Par ailleurs, un même individu peut se développer en forme grégaire ou en forme solitaire en fonction des signaux de son environnement pendant son développement : le patrimoine génétique de chaque individu contient toutes les informations pour les deux formes de criquet. On parle de plasticité phénotypique.

3.2 Examen externe
Si vous comparez l’organisation générale externe du criquet avec celle de l'Ecrevisse, quelles ressemblances et dissimilitudes pouvez-vous repérer ?s.
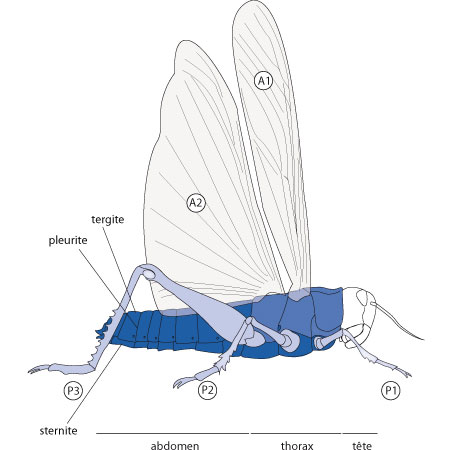
Le corps
On distingue nettement trois régions du corps : la tête, le thorax et l'abdomen. Le corps est métamérisé, et chaque métamère est entouré dorsalement par le tergite, ventralement par le sternite et latéralement par les pleurites.
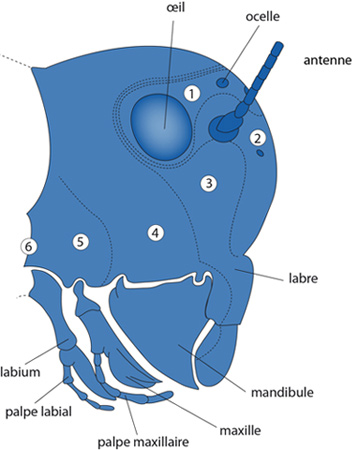
Les antennes
Au sommet de la capsule céphalique, vous distinguez les antennes. Elles sont sensibles au toucher, et bien pourvues de chémorécepteurs. Les deux yeux à facette forment des images; la photo-réception concerne aussi les ocelles, plus petits. Les pièces buccales sont dirigées vers le bas: le labre doté d'une forte musculature maintient les aliments et les pousse entre les autres pièces buccales. Les mandibules saisissent la nourriture. Le labre cache les maxilles, qui sont les pièces masticatrices; on en voit seulement le palpe maxillaire qui dépasse. Ce palpe est constitué de plusieurs articles, et est doté d'une sensibilité tactile et chémoréceptrice. Le labium porte une paire de palpes labiaux, de fonctions semblables à celle des palpes maxillaires. Toutes ces pièces buccales délimitent une cavité que l'on appelle pré-orale. Elle est limitée en avant par le labre, en arrière par le labium, et sur les côtés par les mandibules et maxilles.
Sur le schéma, les limites des métamères sont indiquées en pointillé. Le métamère 1 porte les yeux et ocelles, le 2 porte les antennes, le 3 le labre et le 4 les mandibules, le 5 les maxilles et le 6 le labium.

La tête
Voici une photographie de la tête en vue latérale. Vous y retrouvez les éléments que nous venons de décrire : yeux ocelles, antennes, labre, mandibule, maxille, labium.
La tête de l'insecte est métamérisée, pourtant la métamérisation ne vous apparaît pas au premier coup d'œil. C'est un exemple typique de la tagmose causée par les mouvements complexes des pièces lors de la morphogenèse, qui font s'interpénétrer, se réduire, s'hypertrophier, s'enchevêtrer, se recouper les métamères comme les pièces d'un puzzle. Chez un animal régionalisé, spécialisé comme notre insecte, les métamères ne sauraient plus être assimilés à des tranches de saucisson. Les lignes que l'on aperçoit au binoculaire sur la capsule céphalique des insectes n'ont rien à voir avec les sutures entre métamères. En fait, ces lignes ont un rôle secondaire de renforcement, ou au contraire sont des lignes de moindre résistance le long desquelles le tégument se rompt au moment de la mue. Seules des études morphologiques comparées et morphogénétiques extrêmement poussées et précises sont à même de retrouver les limites véritables entre métamères.
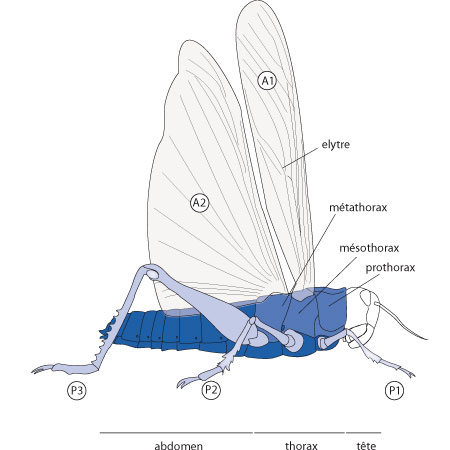
Le thorax
Le thorax du Criquet comprend 3 métamères, appelés respectivement dans le sens antéro-postérieur: prothorax, mésothorax et métathorax. Chacun de ces segments porte une paire de pattes, et les deux derniers portent en outre chacun une paire d'ailes. Les ailes sont insérées entre le tergite et le pleurite. L’origine des ailes n’est pas encore clairement définie. Les insectes les plus primitifs ne possèdent pas d’ailes, ce sont les Aptérygotes. Les insectes ailés sont appelés les Ptérygotes. Une première hypothèse en fait des expansions du tégument, indépendantes d’organes locomoteurs et acquises bien plus tard au cours de l’évolution. Une autre hypothèse les fait dériver d’une transformation complète des organes locomoteurs utilisés pour la marche. Il s’agirait d’une modification du rameau locomoteur ou d’un exopodite et de sa migration vers le haut, au niveau des pleurites. Les ailes antérieures sont appelées élytres chez le Criquet; leur rôle dans le vol est négligeable et se situe surtout dans la stabilisation de l’animal, et la protection des ailes postérieures en les recouvrant quand l'animal est au repos. Les ailes postérieures ont une surface considérable, et le Criquet les replie lorsqu'il atterrit. Ce sont les instruments du vol, et des nervures les renforcent.
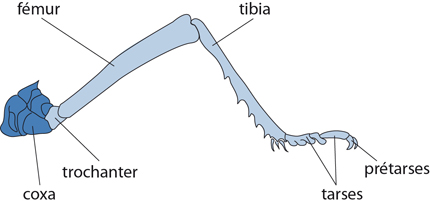
Les pattes
Voyons maintenant les pattes. Les trois paires sont construites sur le même plan, mais leur taille et leur orientation sont variables: la première dirige la marche, le deuxième soutient le corps, et la troisième propulse; chez le Criquet, cette dernière est extrêmement développée et permet à l'animal de sauter. Chacune d'elles comporte à la base un coxa; un trochanter fait l'articulation du coxa avec le fémur; un tibia, un tarse et un pré-tarse muni de griffes. Cet examen externe du thorax démontre bien que cette partie du corps s'est spécialisée dans la fonction de locomotion, que ce soit le vol, le saut ou la marche.

L'abdomen
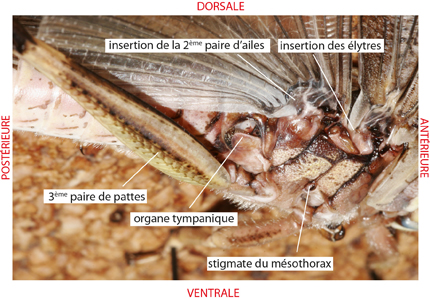
L'abdomen comporte 11 métamères (plus un telson vestigial), mais les 3 derniers chez la femelle, les 2 derniers chez le mâle, sont singulièrement réduits, si bien que la femelle paraît en posséder 8, et le mâle 9. Chaque segment abdominal est percé latéralement d’orifices respiratoires appelés stigmates. Le premier segment abdominal porte sur ses flancs des organes tympaniques sensibles aux sons.
Si la tête s'est surtout spécialisée dans les fonctions de relation et de préhension, et le thorax dans les fonctions de locomotion, l'examen externe de l'animal nous montre que l'abdomen s'est spécialisé dans les fonctions végétatives dont la reproduction.

L'abdomen
Chez la femelle, l'abdomen se prolonge par un ovipositeur, un organe solide qui lui permet d'introduire ses œufs dans le sol. Chez le mâle, des plaques génitales maintiennent la femelle pendant l'accouplement.
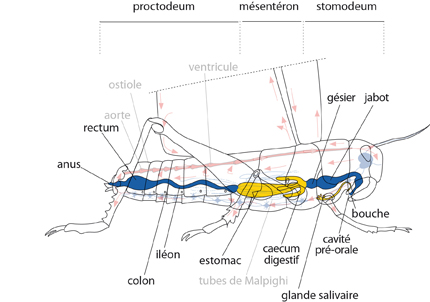
Anatomie interne
Voici un premier schéma, simplifié, de l'anatomie interne du criquet. Il situe les organes principaux. Prenez le temps de le commenter, de le comparer avec ce dont vous vous rappelez de l'Ecrevisse.
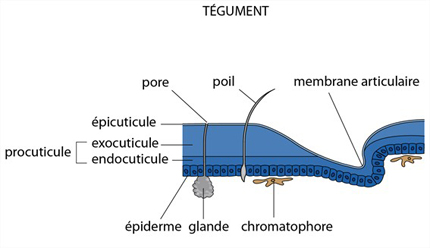
La cutile
L'épiderme secrète la cuticule. Fondamentalement, la structure de la cuticule est semblable à celle de l'Ecrevisse. Avec une nouveauté cependant: des cellules glandulaires sécrètent une pellicule de cire qui recouvre toute la surface. Son épaisseur est minime, mais comme elle est rigoureusement imperméable, elle limite très efficacement la déperdition d'eau. La pellicule de cire est très fragile. Une poudre abrasive, par exemple de l'alumine, de la magnésie ou de la poudre de diatomées, peut constituer un insecticide simple et provoquer une mort rapide de l'insecte par déshydratation, après érosion de la cire. On les mélange parfois aux céréales entreposées dans les silos pour éviter la destruction des stocks par les insectes. Chez les insectes, la cuticule n'est pas minéralisée; cet alourdissement serait bien gênant pour le vol.
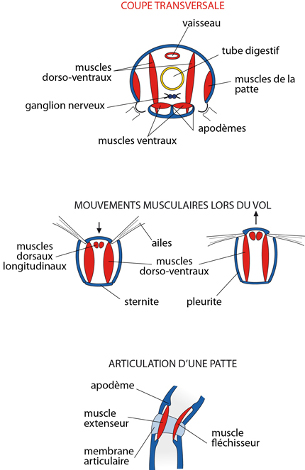
Les muscles
De même, nous ne décrivons pas la musculature de l'insecte, tout entière constituée de muscles striés, y compris ceux qui entourent les viscères. Voici quelques schémas. En a: une illustration des muscles longitudinaux, dorso-ventraux et obliques dans un segment; en b: les élongations et contractions pendant le vol et la position subséquente des ailes; en c: l'articulation d'une patte. Vous êtes capables de commenter ces différents schémas.
3.2.3. Système digestif
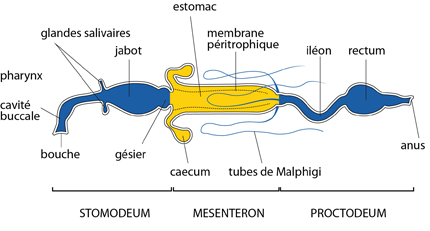
Les criquets sont phytophages: ils consomment des végétaux. Le labre pousse les aliments dans la cavité pré-orale où les maxilles les broient. Dans cette cavité, les glandes salivaires déversent des enzymes dans une solution qui sert à dissoudre les aliments et à lubrifier les pièces buccales. Enfin, les fragments de feuilles mâchés et enduits de salive pénètrent dans le tube digestif. Comme chez l'Ecrevisse, on distingue trois grandes régions le long du tube, en fonction de l'origine embryonnaire de l'épithélium, et d'ailleurs du rôle qu'elles ont: : 1) le stomodeum à revêtement ectodermique, où s'accumulent les aliments et se poursuit le malaxage, 2) le mésentéron à revêtement endodermique, où se poursuit la digestion chimique et où se réalise l'absorption, et le 3) proctodeum à revêtement ectodermique, qui a un rôle d'absorption de l'eau et des sels qui peuvent encore être utilisés par l'organisme avant l'éjection des matières fécales.
Le stomodeum
Détaillons le stomodeum. La nourriture remonte le pharynx et pénètre dans l'œsophage pour déboucher dans le jabot. C'est une poche plissée, logée dans le prothorax. Elle accumule les aliments, et les déverse progressivement dans la poche suivante: un proventricule ou gésier très musclé. L'épithélium du proventricule est armé de dents cuticulaires, qui achèvent la mastication des végétaux.
Le mésentéron
La suite du dessin indique le mésentéron. Ces végétaux réduits en bouillie très partiellement décomposée par les enzymes salivaires, entrent dans le mésentéron, site principal de production enzymatique, de digestion et d'absorption. Dès l'entrée, le mésentéron présente 6 caeca disposés en rosette. Ces évaginations augmentent beaucoup la surface intestinale, puisque chacun mesure 10 mm de long. Ils sont allongés parallèlement au tube digestif, vers l'avant et vers l'arrière. La partie postérieure du mésentéron porte d'autres tubules aveugles: ce sont les tubes de Malpighi, qui représentent le système excréteur. Une particularité des Insectes: une membrane appelée péritrophique est superposée à l'épithélium du mésentéron; c'est une couche très mince d'une sorte de chitine fort particulière, élaborée par les cellules du stomodeum.
Le proctodeum
Le proctodeum comporte trois régions successives appelées un peu abusivement: iléon, colon et rectum, dont l'épithélium est couvert de cuticule. Le rectum est la partie la plus remarquable: à son niveau, l'épithélium épaissi possède une extraordinaire capacité de récupération de l'eau, de sels et de quelques nutriments résiduels, non encore assimilés par le mésentéron. Enfin, ce qui ne peut vraiment plus être utilisé est éjecté par l'anus.
L'hémolymphe
Vous voyez que sur la surface du mésentéron, les nutriments traversent la membrane péritrophique, puis la paroi intestinale. L'hémolymphe, qui circule librement dans toutes les lacunes et les espaces du corps, se charge de récupérer les nutriments, et de les distribuer aux tissus.

Le vaisseau dorsal
Chez les insectes, le système circulatoire est ouvert et simple. C'est le vaisseau dorsal qui préside à la bonne circulation de l'hémolymphe dans l'organisme; il est rattaché à la paroi du corps latéralement par des muscles. La partie postérieure de ce vaisseau montre sept dilatations, des ventricules contractiles. Chaque ventricule se situe au niveau d'un métamère et est percé de deux ostioles latérales, munies de valves. Ce vaisseau dorsal est étendu dans une cavité: le sinus péricardique, séparé du sinus péri-intestinal par un diaphragme dorsal discontinu. Lorsque le corps se contracte, l'hémolymphe est pulsée dans le sinus péricardique, elle pénètre dans le vaisseau par les ostioles. Les contractions des ventricules et de l'aorte qui les prolongent vers l'avant propulsent l'hémolymphe dans la tête, d'où elle passe à nouveau dans tout le corps, y compris dans les appendices, en suivant un trajet déterminé par d'autres diaphragmes. Contrairement à l’écrevisse, il n’y a pas d’autre vaisseau.
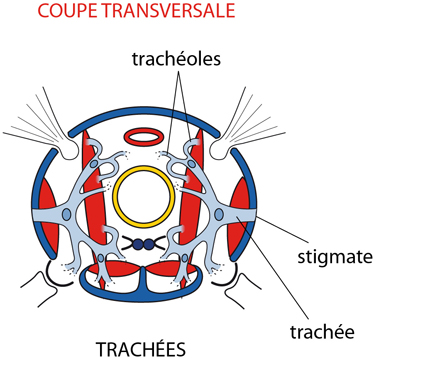
3.2.4. Système respiratoire
Les insectes disposent d’une innovation majeure qui leur a permis de s’adapter à la vie terrestre : le système respiratoire est découplé du système circulatoire. Les échanges respiratoires s'opèrent avec l'air atmosphérique par un système extrêmement ramifié de stigmates, trachées et trachéoles. Les plus petites divisions des trachées, les trachéoles, ont un diamètre de moins d'un micron. Ces minces tubes cuticulaires se ramifient d'une trachée, puis une nouvelle fois encore à la surface des cellules dans les organes internes. La cuticule des trachéoles n'est pas évacuée à la mue comme celle des trachées; après la mue, les nouvelles trachées sont unies aux anciennes trachéoles par une sorte de colle de composition inconnue.
Dans le système trachéen, le transport des gaz se fait par diffusion le long d'un gradient de concentration, aidé par une ventilation. Cette ventilation résulte des mouvements du corps, de l'activité musculaire, qui compriment les sacs aériens. Au bout, dans les trachéoles, l'oxygène et le CO2 sont échangés par diffusion simple. Une bonne partie du CO2 est évacuée aussi par la surface corporelle. Au contraire des trachées, les trachéoles sont perméables à l'eau, et leur extrémité est emplie de liquide intervenant dans le transport final des gaz, qui y entrent en solution. Vous repérez ce liquide, en bleu sombre sur le dessin, au bout des trachéoles. On a pu montrer que le niveau de ce liquide augmente ou diminue en fonction de la pression osmotique des tissus voisins.
Vous pouvez vous imaginer que pendant les vols migratoires de longue durée, les mouvements sont très intenses, les rythmes de contraction et relâchement des muscles sont rapides et soutenus et les échanges gazeux très importants.
L'oxygène brûle les réserves du corps pour les transformer en énergie. Les réserves de glycogène et de graisse sont stockées dans des cellules graisseuses logées en nappes çà et là dans la cavité générale du corps. Ces réserves doivent être abondantes, si l'on songe qu'en une heure de vol, le criquet perd presqu'un pour cent de son poids, et qu'il peut assumer des vols de 9 heures consécutives.
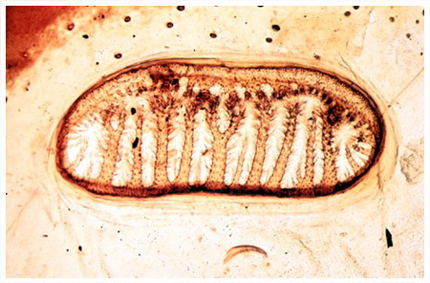
Les stigmates
Les orifices respiratoires, ce sont les stigmates que nous avions repérés lors de l'examen externe. En voici une préparation microscopique. Les stigmates sont pourvus d'un appareillage de fermeture et de filtration, constitué de valves et de poils, qui s'oppose à la déperdition d'eau, l'entrée de poussières et de parasites. L'ouverture et la fermeture des stigmates sont contrôlées par un jeu de muscles innervés directement, et la fréquence des ouvertures est liée à la tension d'oxygène et de CO2 dans l'hémolymphe. Les stigmates donnent accès aux trachées.

Les trachées
Les trachées sont des invaginations de l'ectoderme colorées en rose sur la photo suivante ; on ne s'étonnera donc pas de voir leur lumière tapissée de cuticule. Les plus grosses sont renforcées par un épaississement en spirale très serrée de cette cuticule, que vous apercevez sur la photo ; les plus petites sont renforcées par des anneaux. Les anneaux résistent à la compression, c-à-d qu'ils préviennent le collapsus, mais permettent au tube de s'étirer. A certains endroits, les trachées se dilatent en sacs aériens, tapissés seulement d'une mince cuticule, et donc particulièrement déformables.
3.2.6. Système excréteur
L'hémolymphe a une grande importance physiologique, puisqu'elle charrie les nutriments et les déchets du catabolisme, mais elle n'a qu'un rôle respiratoire très réduit, contrairement au sang des autres animaux que nous avons examinés. Chez le Criquet comme chez les autres Insectes, les échanges respiratoires se font directement entre les trachéoles et les cellules. Il n’y a donc généralement pas de pigments sanguins pour le transport de l’oxygène.
Dans la cavité générale du corps, l'hémolymphe chargée de déchets les déverse dans les tubes de Malpighi, insérés à la jonction mésentéron-proctodeum. Le produit terminal principal du catabolisme azoté est l'acide urique; il est prélevé par le tube de Malpighi mélangé avec de l'eau, des acides aminés, des sucres, des sels. Ces dernières substances regagnent en partie l'hémolymphe à partir des tubes de Malpighi. Puis l'urine provisoire est déversée dans le proctodeum. En profitant des fortes capacités de réabsorption du rectum, l'eau, les sels, les sucres et les acides aminés regagnent l'hémolymphe. L'acide urique précipite sous forme de cristaux, évacués avec les excréments par l'anus.
Il ne faudrait pas croire que toute l'excrétion s'opère par les tubes de Malpighi. Les processus sont bien plus variés. Des cellules phagocytaires réparties dans l'organisme, et rassemblées surtout près du cœur prélèvent les détritus complexes et les dégradent. On trouve aussi des phagocytes dans le tissu graisseux. Ils servent à stocker l'acide urique. Les sels en excès et d'autres substances sont déposés dans la cuticule, pour être éliminés à la prochaine mue. Mais les tubes de Malpighi sont « l'organe excréteur » défini.
3.2.7. Système nerveux
En-dehors des yeux, ocelles et organes tympaniques, les organes des sens sont essentiellement concentrés sur les appendices. Les chémorécepteurs sont abondants sur les antennes, les pièces buccales, et les pattes. Les chémorécepteurs olfactifs sont insérés dans la cuticule, tandis que les chémorécepteurs gustatifs sont portés par des poils. Les terminaisons sensorielles parviennent à la surface de la cuticule par de fins canalicules. La pression, les vibrations, et des changements de tension de la cuticule sont détectés par des poils rigides dont les mouvements sont perçus par des récepteurs à la base, ou encore grâce à des amincissements de la cuticule sous lesquels se dressent des prolongements sensoriels.
D'autres récepteurs de tension existent notamment au niveau des articulations. Il s’agit d’une portion amincie d’exocuticule dépourvue d’endocuticule qui vibre comme la peau d’un tambour. Le même principe est utilisé par les organes tympaniques logés dans le système respiratoire. Vous en voyez l'orifice sous les ailes, à la base de l'abdomen. Les vibrations sonores viennent frapper la membrane tympanique, qui les transmet à la masse d'air circulant dans le système respiratoire, et ces vibrations sont enregistrées par des cellules sensorielles.

Les ocelles
Le criquet possède deux types de photorécepteurs : les ocelles et les yeux composés. Chacune des trois ocelles du criquet est organisée un peu comme l'ommatidie d'un œil composé: elle peut donc seulement détecter des changements de l'intensité lumineuse, et même être sensible aux très faibles intensités, mais elle est incapable de former une image. Quant aux yeux composés, leur structure ressemble fort à celle décrite chez l'Ecrevisse. Le champ visuel de chaque œil est proche de 180 degrés. Le Criquet perçoit une grande variété de couleurs, mais comme chez nombre d'autres insectes, des expériences ont montré que la sélection dans le prisme est différente de la nôtre: elle est déplacée légèrement vers l'ultraviolet.
Exercice
La vue joue un rôle important dans la transition entre les formes de Criquets solitaires et grégaires. Par quelles expériences tenteriez-vous de démontrer sans ambiguïté cette affirmation? (n'oubliez pas qu'une opération traumatisante sur cette pauvre bestiole perturberait évidemment le mécanisme que vous voulez précisément démontrer).
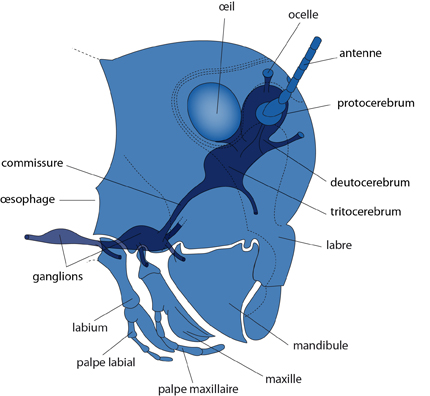
Le système nerveux
Tous ces dispositifs sensoriels sont reliés à un système nerveux, bâti sur un plan très comparable à celui que nous avons décrit chez l'Ecrevisse. Les nerfs en provenance des yeux pénètrent dans le protocerebrum, qui renferme les centres optiques. Le deutocerebrum reçoit les nerfs de la première paire d'antennes (en lien avec l’olfaction) et contient leurs centres d'association. Le tritocerebrum reçoit les nerfs de la seconde paire d'antennes et innerve la bouche ainsi qu'une partie du tube digestif (le centre tactile). Connectées à la commissure du tritocerebrum, en arrière de l'œsophage, nous trouvons la chaîne nerveuse ventrale, médiane et formée de ganglions métamériques plus ou moins fusionnés. Ces ganglions innervent notamment les appendices buccaux, thoraciques et abdominaux.

Système nerveux
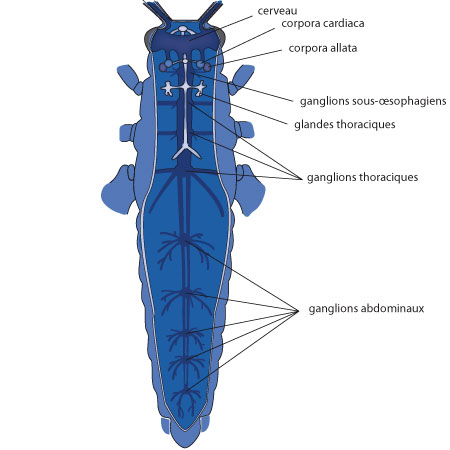
3.2.8. Système endocrine
Comme chez l'écrevisse, ce sont les hormones qui règlent notamment la croissance et les mues. L'emplacement des organes producteurs d'hormones, étroitement associé au système nerveux: les corpora cardiaca, les corpora allata et les glandes thoraciques.
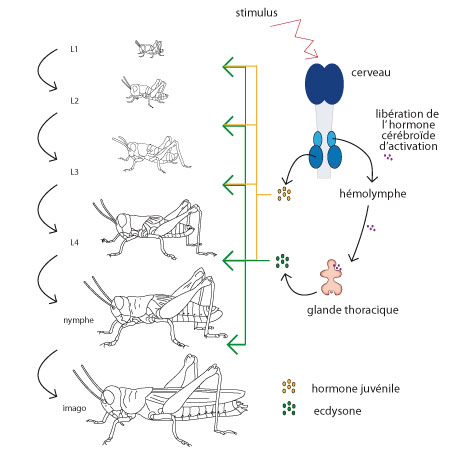
L'ecdysone
Après intégration des stimuli externes et internes dans le cerveau, l'information est transmise à certains groupes de neurones neurosécréteurs qui élaborent des hormones et les véhiculent le long des axones vers des organes de stockages appliqués à la surface du cerveau: les corpora cardiaca. Libérées dans l'hémolymphe, ces neurosécrétions vont stimuler les glandes thoraciques, qui à leur tour secrètent l'hormone de mue : l'ecdysone.
L'ecdysone est libérée dans l'hémolymphe, elle parvient aux tissus et y stimule la croissance et la mue.
Mais pendant les stades nymphaux, une autre neurosécrétion stockée dans les corpora cardiaca excite les corpora allata voisins. En réaction, ceux-ci sécrètent l'hormone juvénile, qui agira aussi sur les tissus. Comme son nom l'indique, l'hormone juvénile est responsable du maintien des caractères jeunes.
C'est donc surtout le jeu de deux hormones: ecdysone et hormone juvénile qui règle le développement de l'insecte. Si les deux hormones sont en concentration suffisante, le résultat sera une mue nymphale. Si l'hormone juvénile n'est plus guère sécrétée, la mue sera imaginale. Et enfin, chez l'imago, les glandes thoraciques sécrétrices d'ecdysone dégénèrent: il n'y aura donc plus de mue, l'animal a son squelette définitif.
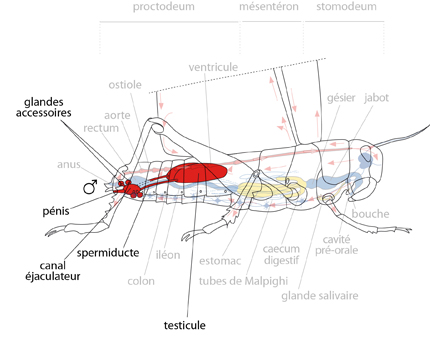
Le système génital du mâle
Le système génital du mâle ou de la femelle occupe une position semblable dans l'abdomen : sous le diaphragme dorsal, au-dessus du tube digestif.
Chez le mâle, les deux testicules accolés produisent les spermatozoïdes et les déversent le long de deux spermiductes, vers un canal éjaculateur unique. Dans ce canal se déversent aussi les sécrétions de deux glandes accessoires qui agglutinent le sperme. Lors de l’accouplement, le sperme transite par le pénis qui fait protrusion et est introduit dans l'atrium génital de la femelle.
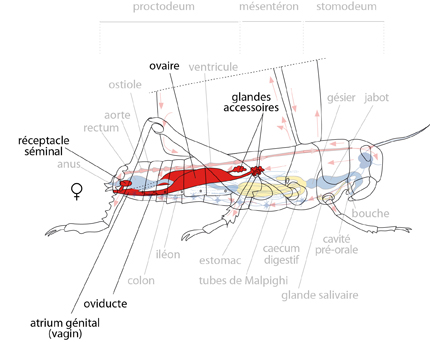
Le système génital de la femelle
Chez la femelle, les ovules se forment dans les ovaires. A maturité, ils passent dans les oviductes. Les glandes accessoires y déversent une sécrétion, qui entoure les ovules et durcira au contact de l'air, constituant une oothèque. Les ovules sont conduits dans l'atrium génital où la fécondation se produit. Cette fécondation survient lors de l'accouplement, ou longtemps après! Comment? Observez le réceptacle séminal: cette poche peut stocker des réserves de spermatozoïdes, et les entretenir pour qu'ils survivent plusieurs mois. Aussi des ovules arrivés à maturité longtemps après l'accouplement pourront quand même être fécondés. Ce phénomène est très favorable à la fécondité de l'espèce, parce que la fertilité de la femelle est influencée par des facteurs tels que la température, l'humidité, les ressources alimentaires. De plus, les criquets en phase solitaire mènent une vie très individuelle. Si la femelle rencontre un mâle en période non fertile, la provision de spermatozoïdes qu'elle reçoit généreusement pourra féconder les ovules qu'elle produira plus tard, lorsque les conditions extérieures seront plus favorables, même si elle ne trouve plus de mâles à ce moment-là.
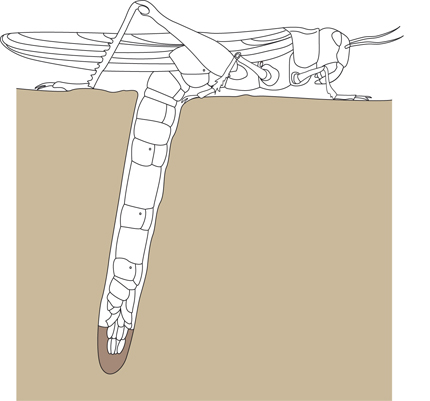
Le système génital de la femelle
La femelle pond ses œufs profondément dans le sol où ils sont protégés des températures extrêmes. Grâce à son ovipositeur très dur, elle perce un trou, et plus elle creuse, plus son abdomen s'allonge. En une seule ponte, elle dépose une centaine d'œufs, et elle peut pondre plusieurs fois par an. Au bout de deux semaines, chaque œuf aura produit une nymphe qui ressemble grosso modo à l'adulte, et le petit criquet remonte à la surface.
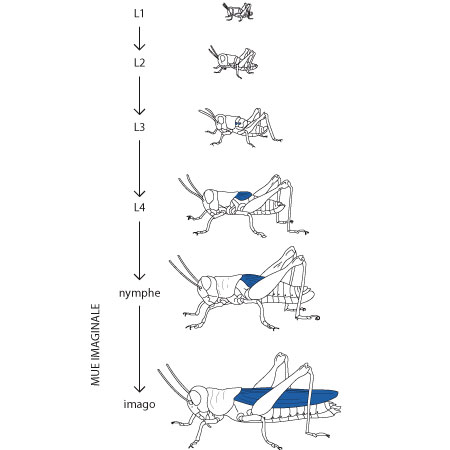
La nymphe
La nymphe grandit par mues successives. Au cours de la croissance et des mues qui lui sont associées, l'animal acquiert de nouvelles structures qui le font ressembler de plus en plus à l'adulte. Par exemple, vous voyez sur ce schéma à quel moment les ailes apparaissent. De même le système génital n'arrivera à maturité fonctionnelle qu'au moment de la dernière mue. On appelle « nymphe » l'individu dans sa phase de développement et « imago » la forme adulte de l'insecte parfait. Chaque mue est nommée «mue nymphale» sauf la dernière dite «mue imaginale» parce qu'elle produit l'imago. Ce développement peut durer de 30 jours à plusieurs mois en fonction de la température. Les criquets font partie du groupe des Hétérométabole car la première larve ressemble déjà à l’adulte. Contrairement aux papillons qui appartiennent aux Holométaboles et où le dernier stade larvaire subit une métamorphose complexe pour se transformer de chenille en papillon.

4. Exemple type 3 : L’Epeire diadème Epeira diademata (Chélicérate)
Nous changeons de groupe d’Euarthropode pour nous intéresser aux Chélicérates. Le dernier exemple d'Euarthropode que nous étudierons donc en profondeur, ce sera une Araignée très commune en Europe: l'Epeire diadème ou Epeira diademata, vulgairement appelée araignée porte-croix, à cause des taches claires qui dessinent une croix sur la livrée plus sombre.
Exercice
En examinant cette image, quelles différences majeures lui trouvez-vous par rapport au Criquet et à l'Ecrevisse ? Prenez un peu de temps pour réfléchir et noter vos observations.
Ici, l'Epeire est accrochée sur sa toile, dans l'attente patiente qu’une proie se prenne à son piège. Ses proies, ce sont des mouches, des moustiques et d'autres insectes dont l’araignée se nourrit en quantité. La cuticule chitineuse qui recouvre le corps est enduite d'une huile qui évite l’adhésion à la toile. Notez aussi les nombreux poils sensoriels.

4.1 Examen externe
Le corps de l’araignée diadème apparaît composé de deux parties bien distinctes : une petite et l'autre beaucoup plus grande ; la petite est le céphalothorax (ou prosome) sur lequel s'articulent les pattes; l'abdomen (ou opisthosome) est dépourvu d'appendices locomoteurs. Le céphalothorax porte les yeux et 6 paires d'appendices au total. Les yeux sont au nombre de 8, disposés sur deux rangs parallèles.

L'araignée'
Contrairement à l'écrevisse et au criquet, l'araignée ne dispose pas d'yeux composés mais bien d‘ocelles de structure interne complexe, surtout chez des espèces chasseresses où la vue est importante pour poursuivre les proies.

Les chélicères
Comme indiqué ici, la bouche ventrale est encadrée de 2 paires d'appendices, les chélicères et les pédipalpes. Les chélicères sont les pièces buccales typiques des Araignées. Chaque chélicère se compose d’une tige mobile articulée au céphalothorax et d’un crochet dur.

L'Epeire
Quand l'araignée attaque une proie prise au piège dans sa toile, elle ouvre ses crochets, écarte au maximum ses chélicères l'une de l'autre, puis elle les resserre dans le corps de sa victime et lui injecte une dose de poison qui la paralyse aussitôt. Chez l'homme, la morsure de l'Epeire ne provoque qu'une réaction superficielle sans gravité. C’est le cas de la majorité des araignées, certaines ne parviennent même pas à percer la peau humaine. Cependant, une minorité d’entre elles peuvent provoquer des piqûres douloureuses et dans quelques rares cas, la mort: pensez à la Veuve noire! En plus du venin, le canal interne aux chélicères laisse passer une sécrétion salivaire enzymatique puissante, qui provoque une liquéfaction rapide des tissus.
Les pédipalpes situés en arrière des chélicères se composent aussi de deux éléments: une large pièce masticatrice et un palpe articulé. Les palpes sont sensibles au toucher et garnis de chémorécepteurs. Ils remplissent le rôle joué par les antennes chez les Insectes. Chez l'araignée mâle, ces palpes sont modifiés en organe copulateur.
La Locuste
Les pattes locomotrices, au nombre de 4 paires comme chacun le sait, comptent les 6 articles décrits chez la Locuste, plus une pièce intermédiaire. Enumérons: le coxa, le trochanter, le fémur, puis la patelle qui est cette pièce intermédiaire, et enfin, le tibia, le tarse et le prétarse muni de griffes.
Au niveau de l’abdomen, deux stigmates respiratoires s'ouvrent à la partie antérieure, et très près de ceux-ci l'orifice génital. Juste en avant de l'anus terminal, il y a 6 filières disposées en rosace (vous les voyez sur la figure de l’examen interne). Ces excroissances sont mobiles et l'araignée les utilise avec dextérité lorsqu'elle construit sa toile. La soie contient de la kératine et est sécrétée par de nombreuses glandes dont les canaux débouchent au sommet des filières. Ces sécrétions ne sont pas toutes du même type, et l'araignée choisit judicieusement quelle sorte de soie elle sécrète lors de son travail: plus résistante, plus gluante. D'abord liquide, la soie se solidifie au premier contact de l'air. La toile sert de filet de chasse. Parfois même, l'araignée ligote sa proie récalcitrante dans un bel emballage de soie pour mieux l'immobiliser. A l'époque de la ponte, la femelle tisse un cocon très épais dans lequel elle déposera quelques 600 œufs qui s'y développeront à l'abri des rigueurs de l'hiver. Les jeunes sortiront au printemps, subiront plusieurs mues, deviendront adultes en été et s'accoupleront à l'automne.

4.1 Examen interne
Voyons d’abord le système digestif. La nourriture liquéfiée est pompée, aspirée par le pharynx, l'œsophage, et l'estomac, puis passe dans le mésentéron absorbant. Le mésentéron prend énormément de place avec tous ses diverticules qui s'allongent jusque dans les pattes et remplissent une bonne partie de l'abdomen. Le proctodeum conduit les détritus à l'anus.
L'air pénétrant par les stigmates parvient dans deux invaginations ectodermiques renforcées de cuticule et disposées en lamelles: ce sont les poumons feuilletés ou poumons lamellaires. Entre les lamelles emplies d'air circule l'hémolymphe qui s'oxygène, largue son CO2 et retourne dans la cavité péricardique entourant le cœur dorsal, percé d'ostioles.
Il vous reste encore à localiser le système génital, nous avons représenté une femelle ici, et l'appareil excréteur: deux tubes de Malpighi raccordés à la jonction mésentéron-proctodeum.
Enfin, le système nerveux en bleu, est hyperconcentré dans le céphalothorax: un gros cerveau antérieur, relié à une masse ventrale de ganglions fusionnés par deux commissures autour du pharynx. Attention, chez les chélicérates le cerveau est en 2 parties et le deutocérébrum est absent.
Voilà, ce fut bref pour l'Araignée, mais vous disposez maintenant d'un représentant des trois lignées principales d’Euarthropodes, que vous pouvez comparer à loisir. Maintenant, nous allons aborder l'évolution et la fantastique adaptabilité de ce phylum.

5.1. Origine évolutive des Arthropodes
C’est dans le phylum des Panarthropodes que l’on trouve la majorité des espèces vivantes sur terre. Près d’un million d’espèces ont été décrites au sein de ce groupe, ce qui représente plus de 85 % de toutes les espèces connues. Il y en aurait bien plus encore à découvrir puisque l’on estime le nombre d’espèces de Panarthropodes dans une fourchette de 5 à 50 millions. Ils ont conquis pratiquement tous les habitats: marin, dulçaquicole et terrestre; ils sont sans doute les organismes qui ont eu le plus de succès sur Terre.
Vu leur abondance et la diversité des formes, la systématique des Panarthropodes est complexe, il ne nous sera pas possible de décrire tous les groupes. Par contre, nous allons tenter de répondre à nos trois questions de base et nous allons donner une vision la plus synthétique possible de ce que l’on connaît de leur positionnement phylogénétique actuel.
Les premiers Arthropodes
Les gisements fossiles du schiste de Burgess en Colombie Britannique, de Chengjiang du sud de la Chine et de Sirius Passet dans le nord du Groenland offrent des informations sur l’évolution des PanArthropodes. Les premiers Arthropodes incontestables apparaissent au Cambrien inférieur, à -570 millions d'années environ, au début de l'ère primaire.
Les découvertes du schiste de Burgess en Colombie Britannique montrent qu’il y a 540 millions d’années, les formes d'Arthropodes foisonnent et sont déjà étonnamment complexes et spécialisées. C’est ce que l’on appelle l’explosion du Cambrien, c’est-à-dire une diversification surprenante des formes de vie qui s’est déroulée sur un laps de temps relativement court par rapport au temps géologique. Cette diversité des plans fondamentaux d’organisation du vivant s’appelle : la disparité. Peut-être est-ce le résultat de l'absence de compétition qui devait exister avant que toutes les niches écologiques des mers du Cambrien ne soient occupées? A ce moment, toute forme nouvelle avait une chance de se maintenir. D’autres hypothèses ont également été émises.
Des Anomalocarididea
La plupart de ces premiers Arthropodes se sont rapidement éteints, sans doute parce que les animaux les moins bien adaptés ont été remplacés par d'autres mieux adaptés, sans doute aussi parce que certains ont été moins «chanceux»; ainsi, le nombre de morphologies de base, fondamentalement différentes, a-t-il progressivement diminué. Au cours de l’évolution, on assiste donc à une diminution de la disparité et à une augmentation de la diversité au sein des plans d’organisation qui se sont maintenus. Il semble que le groupe souche des Arthropodes pourrait être celui des Anomalocarididea (avec les fossiles de Anomalocaris et Hurdia) et celui des Opabinia qui disposent déjà d’yeux à facette et pédonculés mais pas encore d’un exosquelette sclérifié. Néanmoins, les relations phylogénétiques au sein des groupes fossiles restent particulièrement difficiles à établir.
Au Cambrien, deux groupes de fossiles sont déjà clairement morphologiquement distincts: les Crustacés et les Trilobites.
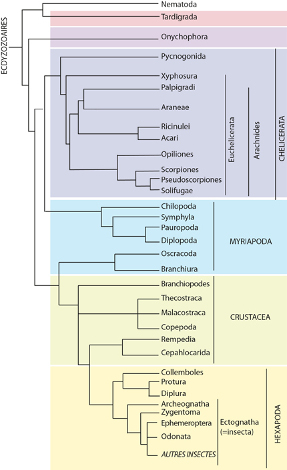
Les Ecdyzozoaires
Sur base du séquençage de l’ARN ribosomique 18S, on a pu montrer pour la première fois que les Arthropodes se rapprochent des Nématodes et d’autres animaux caractérisés par l’existence d’au moins une mue (Ecdysis) au cours du développement. Cette nouvelle façon de voir les relations phylogénétiques sera ensuite confirmée par d’autres études moléculaires sur d’autres marqueurs moléculaires. Au sein des Ecdyzozoaires (les animaux qui muent), les Arthropodes appartiennent à un groupe plus large nommé Panarthropodes qui regroupe les Onychophores, les Tardigrades et les Euarthropodes. Ces groupes ont tous la caractéristique de disposer d’appendices pairs segmentés et placés ventro-latéralement sur chaque segment. Cette caractéristique commune plaide donc en faveur d’un ancêtre commun pour ces groupes, même si les relations exactes de parentés entre les trois clades restent mal définies.

Les Myriapodes
Au sein des Euarthropodes, la plupart des auteurs sont maintenant d’accord sur la monophylie des espèces possédant des mandibules et rassemblées dans le groupe des Mandibulates, alors que les espèces possédant des chélicères forment le clade des Chélicérates. Les Myriapodes sont également considérés comme un groupe monophylétique que cela soit sur le plan des analyses moléculaires ou sur le plan morphologique.
Un petit groupe reste difficile à placer, il s’agit des Pycnogonides, qui se rattachent néanmoins aux Chélicérates et forment avec eux les Chélicériformes. Ils ne possèdent pas de chélicères mais des chélifores. On a tout d’abord cru que les chélifores était innervés par le protocérébrum et non pas par le deutocérébrum comme c’est le cas pour les chélicères. En conséquence, certains auteurs estimaient qu’il pourrait s’agir du « grand appendice » que l’on retrouve chez certains arthropodes primitifs du Cambrien, ce qui en ferait la lignée la plus ancienne du clade des Euarthropodes. Cependant des données plus récentes sur l’expression des gènes Hox montrent clairement que Chélicères et Chélifores sont homologues et liés au segment du deutérocérébrum, même si le deutérocérébrum a ensuite disparu au sein de ce groupe. Les Pancrustacés ou Tetraconota possèdent en commun (comme le nom Tetraconata l’indique) quatre cellules coniques qui constituent le cristallin à l’intérieur des ommatidies. Ils partagent également des caractéristiques au niveau de la neurogenèse, du cerveau et du lobe optique. Les relations exactes entre les différents groupes restent difficiles à ancrer comme le montre l’arbre phylogénétique non enraciné.
5.2 Diversité et relation entre les Groupes
Dans cette section, nous allons décrire les clades majeurs et la diversité qu’ils contiennent. L’arbre nous aide à situer les groupes taxonomiques les uns par rapport aux autres.
Cette section est à lire, mais pas à retenir. Elle illustre la grande diversité d’Arthropodes sur terre.
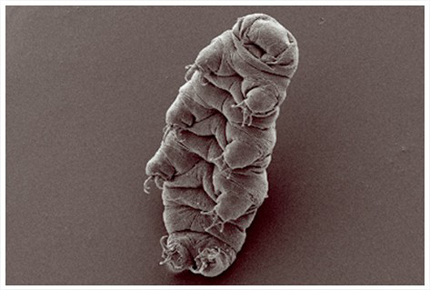
5.2.1 Les Tardigrades
Il s’agit de petits organismes de 0,1 à 1,5 mm. Leur corps est composé de 4 segments, chacun portant une paire d’appendices locomoteurs. Ils sont inféodés à l’eau et vivent dans les substrats humides, particulièrement les mousses et les lichens et on en dénombre à peu près 1000 espèces décrites à ce jour. Ils ont colonisé tous types de milieux y compris les plus extrêmes puisqu’on les trouve en Himalaya jusqu'à 6000 m d’altitude, en Antarctique, dans les eaux profondes jusqu’à 4000 m de profondeur mais aussi dans la mousse qui recouvre nos toitures. Ils sont caractérisés par des adaptations physiologiques extrêmes que l’on appelle cryptobiose. Ainsi, on les a trouvé dans certains déserts où ils peuvent perdre jusqu’à 99 % de leur eau et rester dans un état de complet repos métabolique, semble-t-il pendant des dizaines d’années. Ils montrent aussi une résistance exceptionnelle au froid, à la chaleur et aux rayons X.

5.2.2 Les Onychophores
Il existe 80 espèces d’Onychophores (porteur d’ongle), toutes vivent en région tropicale. Il existe deux familles, les Peripatidae (voir photo) et les Peripatopsidae qui diffèrent par répartition géographique. Ce sont des animaux terrestres porteurs de nombreuses paires de pattes (de 14 à 43), d’une paire d’antennes munies d’yeux à la base. Animal étrange des litières forestières, voici un Péripate. Son nom signifie «qui déambule», même racine que pour la dénomination de certaines dames qui arpentent les trottoirs. Dans les forêts tropicales humides, si on fouille la masse des feuilles en décomposition, on peut trouver ce petit animal long de 5-6 centimètres, à peau veloutée, avec de petits yeux et de longues antennes, et de nombreuses pattes en forme de moignons se terminant par des griffes.

5.2.3 Les Euartrhopodes - 5.2.3.1 Les trilobites
Présents dès le tout début du Cambrien, le groupe de Trilobites a très vite évolué et va ensuite conserver sa morphologie avec peu de modification pendant près de 300 millions d’années avant de disparaître définitivement au Permien, il y a – 248 millions d’années. Peut-être que très adapté, il était peu adaptable aux changements majeurs de cette époque.
La plupart des pièces fossiles des Trilobites ne représentent pas l'animal lui-même, mais sa carapace, abandonnée lors d'une mue. Les spécimens complets sont plus rares.
Voici un Trilobite fossile. Par leur taille, les Trilobites varient de 1 à 75 centimètres de long. Leur corps est large et aplati. Comme chez les autres Arthropodes, une demi-douzaine de segments antérieurs ont fusionné pour former une tête recouverte par un large bouclier; en arrière de ce bouclier, cependant, on ne note aucune tendance à la formation d'un thorax. Chaque segment est articulé à ses voisins et mobile, à l'exception d'un nombre variable de segments postérieurs qui peuvent se souder pour former un pygidium, une queue obtuse. Le corps est très flexible, au point que l'animal entier pouvait s'enrouler en boule pour se protéger. Chaque segment du corps est divisé transversalement en trois parties, d'où le nom de «Trilobite». La zone médiane contient les organes essentiels de l'animal; les deux ailes latérales protègent les appendices qu'elles recouvrent.
Les Trilobites
Sur la face dorsale de la tête, on voit une paire d'yeux composés bien développés. La bouche et les antennes sont reléguées sous le bouclier céphalique. A l'exception des antennes, tous les appendices sont construits sur le même plan : chacun comprend une branchie, sorte de lamelle filamenteuse pour la respiration, et une patte segmentée dont la partie basale, le coxa, est munie de piquants dirigés vers l'intérieur. Ils servent à broyer la nourriture et à la transporter progressivement vers la bouche, de l'arrière vers l'avant. C'est un fait très primitif pour un Arthropode que les appendices céphaliques soient tout à fait semblables à ceux qui leur sont postérieurs et très peu modifiés pour la mastication.
La majorité des Trilobites n'étaient pas des nageurs actifs, mais bien des animaux rampant sur les fonds boueux ou y fouissant à la recherche de nourriture.
Les Trilobites sont les mieux connus des premiers Arthropodes, mais ils ne sont en aucun cas les ancêtres d'un autre groupe, en dépit de leurs caractères primitifs comme l'absence de pattes-mâchoires spécialisées. Le corps très large muni de lobes latéraux est sûrement une spécialisation, même si elle fut précoce et pour un temps couronnée de succès.

5.2.3.2. Les Chélicériformes
Outre la division du corps en prosoma et opisthosoma, la grande caractéristique de ce clade est la possession de Chélicères qui sont homologues de la deuxième paire d’antennes chez les Mandibulates. La deuxième paire d’appendices est appelée pédipalpe et peut être plus ou moins transformée. Autre particularité, le deutérocérébrum a disparu dans ce clade.
Les Chélicériformes sont composé d’une part des Pycnogonides et d’autre part des Euchélicérates. Les relations phylogénétiques sont représentées dans la figure. Nous allons faire une brève description des groupes principaux.

Les Pycnogonides
Les Pycnogonides, parfois appelés araignées de mer possèdent un proboscis proéminent et sont caractérisés par une extrême réduction de l’abdomen, ce qui les rend aisément reconnaissables. Il n’y a ni organe d’excrétion, ni organe de respiration, ce qui est compensé par un très grand rapport en surface sur volume du corps. Une particularité du groupe, les mâles recueillent les ovules fécondés sur leurs ovigères jusqu’à l’éclosion. Il s’agit d’une forme de soins parentaux exercée par les mâles. Connaissez-vous des exemples dans d’autres groupes animaux ? On connaît près de 1300 espèces de Pycnogonides ayant colonisé toute une série d’habitats marins même s’ils restent discrets et relativement peu abondants. Il en existe par exemple sur les côtes de la mer du Nord.

Les Mérostomes
Si l’on devait schématiser l’organisation générale d'un Euchélicérate primitif et rassembler les caractères qu’ils possèdent en commun, cela ressemblerait à ce schéma. C’est le groupe des Mérostomes ou Xiphosure (Limules) qui reste le plus proche de cette organisation.
Tous les Chélicérates ont des pinces appelées chélicères, quelquefois de petite taille, parfois puissantes, juste au niveau de la bouche. La partie antérieure du corps est soudée et comprend la tête et le thorax porteur des appendices locomoteurs, mais il subsiste une longueur considérable du tronc en arrière de cette région céphalothoracique, qui contient une grande partie des organes internes. A l'origine, cette région était segmentée, mais chez tous les représentants évolués des Mérostomes et des Arachnides, ces segments peuvent se souder pour former une structure abdominale unique. Enfin, chez les Euchélicérates, les appendices postérieurs ne sont présents que sous une forme très modifiée, notamment des branchies ou des organes jouant le rôle de poumons.

Les Xiphosures
Les Xiphosures sont le plus ancien groupe de la classe des Mérostomes, et celui qui a duré le plus longtemps. Leur nom signifie «dont la queue est une épée». La Limule que vous voyez appartient à l’une des 7 espèces existant encore actuellement. De nos jours, on les trouve sur le sable, le long des côtes atlantiques de l'Amérique du Nord et le long des côtes pacifiques de l'Asie où elles viennent pondre leurs œufs lors des marées d’équinoxe. Elles peuvent atteindre une cinquantaine de centimètres.
La carapace céphalothoracique, bouclier large et arqué, évoque un fer à cheval épaissi pour protéger une bonne partie du corps. Les pattes, la bouche et les pièces buccales sont entièrement cachées par ce bouclier. Sur la face dorsale, celui-ci ne montre qu'une paire d'yeux composés bien développés. En arrière, la Limule présente une organisation différant beaucoup des conditions supposées primitives: tous les segments abdominaux sont en effet soudés en une masse compacte ne montrant dorsalement aucun indice de sa nature segmentée, sauf une série de petites épines le long de chaque bord. Le corps se termine par une longue et puissante épée caudale.
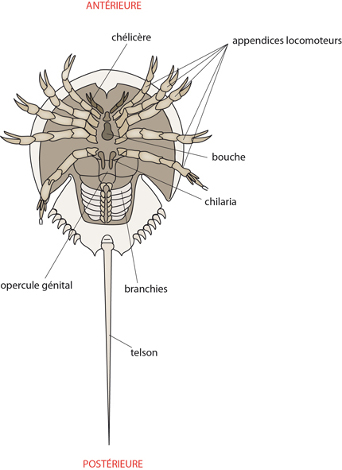
La Limule
Si vous attrapez une Limule par la queue et la retournez, vous pouvez étudier à loisir sa face ventrale très révélatrice représentée dans le dessin suivant. (L'animal ne peut se retourner sur lui-même qu'à la suite de mouvements acrobatiques vigoureux, en se servant notamment de son épée caudale). Vers l'avant, sous le bouclier céphalo-thoracique, on trouve une paire de petits chélicères situés devant la bouche; plus vers l'arrière, 5 paires de pattes puissantes et une sixième paire d'appendices réduits, les chilaria. Sous l'abdomen, 6 paires de larges plaques qui se recouvrent d'avant en arrière: c'est l'appareil natatoire et respiratoire de l'animal dont les éléments sont comparables à un livre, où la surface externe de la page est lisse, et la surface interne comporte de nombreux replis branchiaux fragiles.

Les Euryptérides
Le deuxième groupe des Mérostomes, ce sont les Euryptérides ou Scorpions de mer, éteints depuis longtemps.
Ils se remarquent au milieu des anciens Invertébrés fossiles par leur grande taille: ils pouvaient atteindre 3 mètres! C'étaient des géants de ces temps lointains. Du milieu marin, ils ont conquis les eaux douces.
Les Euryptérides typiques ont quelque peu l'aspect des Limules les plus anciennes, mais plus allongés. En arrière du céphalothorax étroit, une douzaine de segments abdominaux qui ne se soudent jamais et se terminent en une épée caudale. Les chélicères préhensiles ont une taille très variable; chez les uns, ils sont réduits à des organes minuscules et certaines pattes locomotrices contribuent alors à prendre la nourriture. Chez d'autres, les chélicères sont grands, se développent en appendices préhensiles comparables à ceux d'un Homard, mais qui ne sont pas leurs homologues, puisque les pinces des Crustacés sont des pattes thoraciques modifiées.
Comme chez les Limules, les 5 paires d'appendices locomoteurs des Euryptérides sont restreints au céphalothorax. La structure des branchies abdominales n'est pas connue.

Les Arachnides
Les Arachnides constituent un groupe très diversifié avec des modes de vie très différents qu’ils soient aquatiques ou terrestres. Il existe des espèces prédatrices, d’autres parasites, d’autres encore phytophage. Les pages suivantes illustrent quelques exemples de ce clade.
Les plus vieux Arthropodes terrestres sont probablement des Scorpions du Silurien, et leur forme est voisine de celle des scorpions actuels comme celui de la photo, et de celle des Euryptérides, un groupe ne contenant que des fossiles.
Mais cette ressemblance avec les Euryptérides est en partie superficielle. Par exemple, les grandes pinces que l'on trouve chez les Euryptérides dérivent des chélicères; chez les Scorpions, les chélicères, bien qu'efficaces sont de taille modeste, tandis que les grandes pinces dirigées vers l'avant pour saisir les proies sont des pattes locomotrices modifiées. De plus, les Scorpions n'ont pas d'yeux composés, seulement des ocelles, mais assez performantes. Toutefois à bien des égards, Euryptérides et Scorpions sont bâtis sur le même type: même organisation céphalo-thoracique avec les appendices concentrés à la face ventrale, mais comme la première paire est transformée en pince, le Scorpion ne dispose plus que de 4 paires de pattes locomotrices.
La plupart des organes viscéraux s'étendent jusque dans la première demi-douzaine de segments abdominaux, relativement larges; les segments suivants plus minces forment une queue à l'extrémité de laquelle se trouve l'aiguillon du Scorpion; c'est une épine courte et acérée qui peut injecter une toxine douloureuse, et parfois mortelle.
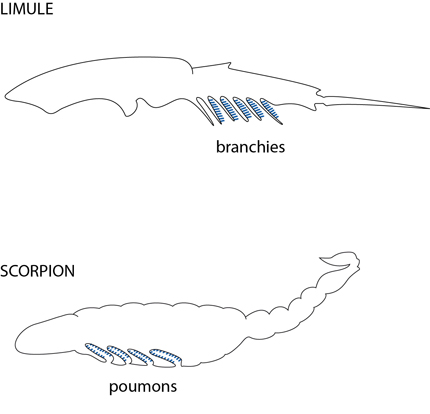
Les Euryptérides
Le passage de l'eau à la terre ferme pose à l'animal de nombreux problèmes fonctionnels. Pour un Arthropode de type Euryptéride, ces problèmes sont moins difficiles à résoudre que pour des animaux dépourvus de carapace chitineuse. La carapace supporte et maintient le corps efficacement dans l'air comme dans l'eau, et pourvu que l'animal ne dépasse pas une trop grande taille, les pattes articulées qui permettraient à l'ancêtre de se déplacer sur le fond des mers ou des étangs, lui permettront de marcher sur la terre ferme. Le problème de la déperdition d'eau par la surface des téguments trouve sa solution avec la mince couche de cire imperméable qui se dépose à la surface de la cuticule. Il reste le problème de la respiration abordé dans le dessin suivant.
Comment absorber l'oxygène de l'air et non plus celui dissous dans l'eau, en évitant en même temps la déperdition d'eau par la surface respiratoire étalée? Les Arachnides ont adopté une solution simple: comme nous l'avons vu, les Limules et les Euryptérides respiraient à l'aide de feuillets branchiaux situés sous l'abdomen. Les tissus délicats de ces branchies fonctionneront encore pour absorber l'oxygène de l'air, pourvu qu'ils puissent rester humides. C'est ce qui s'est passé dans l'évolution des Scorpions: les feuillets branchiaux se sont simplement invaginés à l'intérieur des premiers segments abdominaux, et ils ne s'ouvrent à l'extérieur que par des orifices étroits, les stigmates. Les feuillets branchiaux sont devenus des poumons feuilletés.

Les scorpions
Introduite par la photo suivante, les Araignées que nous connaissons déjà par l'Epeire, sont un autre Ordre. Chez les Scorpions, la première paire de pattes locomotrices était transformée en pinces préhensiles. Chez les autres Arachnides, c'est différent: nous avons vu que les Arachnides ne disposent pas d'antennes véritables. Ces structures sensorielles sont pourtant bien utiles. Les Araignées ont transformé cette première paire de pattes en pédipalpes fonctionnellement comparables aux antennes. Vous les voyez sous la tête, de part et d'autre. Par ailleurs, l'abdomen ne forme plus qu'une seule masse compacte, réunie au céphalothorax par un pédicule étroit. La plupart des Araignées sont inoffensives pour l'Homme et n’arrivent même pas à percer la peau avec leurs chélicères, peu d’espèces sont franchement dangereuses.

Les Faucheux
Vous connaissez tous les Faucheux ou Opilions, ces sortes d'Araignées inoffensives très maladroites sur leurs grandes pattes. En fait, c'est un ordre d'Arachnides où l'abdomen n'est plus réuni au céphalothorax par un pédicule étroit : le corps forme une seule masse globuleuse.

Les Acariens
Enfin, bien que peu connu, les Acariens constituent une classe très importante pour l'Homme, du point de vue économique et de la santé. Les Acariens ont une distribution extrêmement large et ont colonisé tous les milieux y compris les milieux marins et d’eau douce. Ils sont présents aussi bien dans les sources chaudes que dans des eaux à pH 9.6. On les retrouve aussi bien en Régions tempérées ou Tropicales qu’en Antarctique où ils supportent des températures de l’ordre de –60°C ou dans le désert de Namibie à +50 °C.

Les Tétranyques
Cette photo vous montre 2 Tétranyques. Ils appartiennent à l’ordre de Prostigmates et à la famille des Tetranychidae. Ils sont à peine visibles à l'œil nu. Adultes, ils atteignent seulement 600 microns. Les Tétranyques sont posés sur une feuille de haricot et ils ont planté leurs chélicères dans les cellules de l’épiderme dont ils se nourrissent. Les espèces appartenant à ce groupe s’attaquent aux principales plantes cultivées sur terre. En raison de la rapidité de la croissance de leurs populations, ils provoquent des dégâts considérables qui se traduisent en pertes économiques.
Les Gamasides
Ils ont leurs propres prédateurs, les Acariens de l’Ordre des Gamasides couramment utilisés en lutte biologique et dont certains sont produits industriellement et vendus sous marque déposée comme Phytoseiulus persimilis représenté sur cette photo.

Les Acariens des poussières
Certains Acariens apprécient beaucoup notre compagnie; on les trouve dans nos poussières domestiques, dans nos matelas. Ils sont responsables de pas mal d'allergies. Voici un exemple d’Acariens des poussières.

La tique
Ils vont même plus loin beaucoup sont parasites de l'Homme et de ses animaux domestiques. En voici un très commun: une tique de la Famille de Ixodidae. Ces animaux se nourrissent de sang de Vertébrés, car la femelle a besoin de sang pour assurer le développement de ses œufs. On la voit ici dans les poils de son hôte; elle est difficilement reconnaissable. L'abdomen s'est gonflé de sang comme une outre. En passant d'un individu à l'autre, les Tiques peuvent transmettre divers organismes pathogènes.

5.2.3.2. Les mandibulates
Les myriapodes, familièrement appelés Mille-pattes, sont caractérisés par une série d'appendices locomoteurs tous semblables, mais articulés cette fois. Les segments post-céphaliques sont très nombreux, semblables et on ne peut pas distinguer le thorax de l’abdomen. Les Myriapodes sont formés de plusieurs Classes distinctes. Les Chilopodes qui disposent d’une parie de pattes par segment, les Diplopodes, où l’on en compte deux, les Symphyles et les Pauropodes. Ici, les échanges gazeux se font par des trachées. Rappelez-vous quels autres groupes présentes cette particularité ?

Les Chilopodes
Parmi les Chilopodes, nous trouvons par exemple la Forcipule de cette photo, ou encore ce Scolopendre prédateur. Ils ont de nombreux appendices, mais nettement moins qu'une centaine. Ces petits animaux montrent un mélange intéressant de caractères primitifs et de caractères évolués. La tête est petite et globuleuse; vous la voyez ici en photo.
En arrière de la tête, le corps des Chilopodes est extrêmement primitif: à l'exception d'une paire de griffes venimeuses (les forcipules) sur la première paire de pattes, chaque segment est exactement le même que tous les autres et porte une paire de pattes locomotrices. Le dernier porte des gonopodes. Il n'y a aucune régionalisation. Toutefois, ces animaux sont adaptés à la terre ferme: les échanges gazeux s'opèrent par un système trachéen, comme chez le criquet, mais les nombreux stigmates restent continuellement béants.
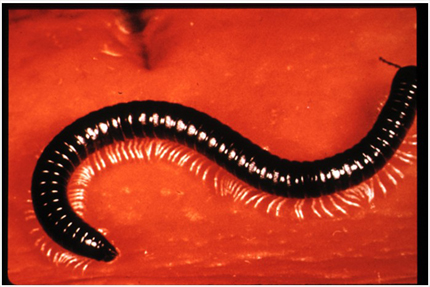
Les Diplopodes
Parmi les Diplopodes, on trouve cette Iule. Les pattes sont encore plus nombreuses, bien que n'approchant pas le millier que leur accorde leur nom populaire. Ce grand nombre de pattes des Diplopodes, comme l'indique leur nom est dû en partie au fait unique chez les Arthropodes, que chaque segment porte deux paires successives d'appendices.
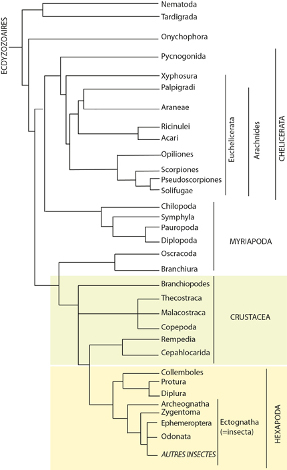
Les Pancrustacés
Les Pancrustacés sont aussi appellé Tetraconata en référence à la présence de 4 cellules responsables de la production du cristallin dans les ommatidies. L’arbre suivant basé sur l’analyse de plusieurs gène nucléaire montre que les insectes (Hexapoda) partage un ancêtre commun avec les Crustacés.
Il existe au plus de 30.000 espèces de Crustacés actuellement décrite. La plupart sont de très petite taille, et d'aspects très hétéroclites, si bien que leur systématique est l'une des plus complexes du monde animal. Nous ne l'aborderons pas complètement ici.
Tous les Crustacés montrent des modifications évolutives nettes par rapport aux Trilobites. Observons la photo suivante. Des appendices antérieurs sont modifiés en organes buccaux efficaces, si bien que les Crustacés les plus grands comme cette Langouste, sont de redoutables prédateurs.

Les Crustacés
Les Crustacés sont les seuls Arthropodes à posséder deux paires d'antennes. Les six métamères et l'acron constituant la tête sont soudés et s'y ajoutent un nombre plus ou moins grand de segments plus postérieurs, allant jusqu'à édifier un céphalothorax. Les segments abdominaux restent articulés. On peut trouver un nombre variable de pattes locomotrices spécialisées, mais il persiste toujours un nombre plus ou moins grand d'appendices biramés caractéristiques.

Les Céphalocarides
La Classe des Crustacés découverte le plus récemment, en 1955 : les Céphalocarides, sont curieusement les plus primitifs des représentants actuels. Ce sont de petits fouisseurs, longs de 4 mm maximum et leur deuxième paire de maxilles ne diffère pas encore des appendices plus postérieurs du tronc.
D'allure presqu'aussi primitive: les Branchiopodes de la photo suivante, petits Crustacés vivant surtout dans les eaux douces, tel Artemia sp., que vous voyez ici. Il est d'élevage très facile. Plus connue, La Daphnie, la puce d'eau si commune dans nos eaux calmes, est enfermée dans une carapace bivalve. C'est aussi un Branchiopode.

Les Branchiopodes
Observons le dessin suivant. Malgré des aspects très divers, tous les membres de cette Classe possèdent comme caractéristique d'avoir des appendices aplatis : l'exopodite et l'endopodite sont des palettes bordées de poils, et l'épipodite, aplati lui aussi, sert de branchie (d'où le nom de la Classe : les Branchiopodes).
La larve Nauplius
Profitons des Branchiopodes pour examiner un élément que nous n'avions pas pu observer chez l'Ecrevisse : la larve Nauplius. C'est la larve de base des Crustacés, et nous l'évoquerons encore par la suite. L'oeil nauplien, tache sombre médiane en avant de la tête, est l'organe photorécepteur.

La larve planctonique
Au sortir de l'oeuf, cette larve planctonique, nageuse, ne possède que trois paires d'appendices: les antennules, les antennes et les mandibules. Les longs poils qui garnissent les deux dernières paires servent à la natation. Un labre impair démesuré se situe en avant de la bouche, à la face ventrale. On ne distingue aucune segmentation externe.

Les Ostracodes
Cette photographie au microscope électronique à balayage montre un minuscule ostracode de 520 millions d'années. La présence d'une carapace est en elle-même une protection pour un petit animal, et les petits Ostracodes, constamment présents par leurs restes fossiles depuis le Cambrien jusqu'à nos jours, ont su en retirer le maximum d'avantages. Vous voyez les appendices articulés et une valve. La carapace durcie s'étend pour former une minuscule coquille bivalve, ressemblant assez fort à celle d'une Moule. Les Ostracodes n'ont pas un grand intérêt évolutif; ils sont cependant utiles au stratigraphe en tant que fossiles repères, notamment pour la prospection pétrolière. Leurs petites coquilles se conservent bien et diffèrent d'une couche géologique à l'autre.

Les Copépodes
Les Copépodes aux antennes hypertrophiées pullulent dans les mers et les eaux douces, cette photo en est un exemple.. Le nombre de Copépodes marins est énorme, la plupart sont planctoniques et se nourrissent de phytoplancton. Ils sont alors un lien principal entre ce phytoplancton et les niveaux trophiques supérieurs, dans la chaîne alimentaire du milieu marin. La nourriture de nombreuses espèces de poissons est constituée en majorité de Copépodes. Ils en deviennent des indicateurs de pêche pour les marins: là où se trouve un banc de Copépodes, les marins sont sûrs de pêcher des harengs.
Les Copépodes ne sont pas seulement bénéfiques: on y rencontre la plupart des Crustacés parasites, externes et internes.

Les Balanes
Ces Balanes, fixées sur des moules appartiennent aux Cirripèdes. Les Cirripèdes se fixent sur n'importe quoi ou presque, et certains trouvent pratique de se faire véhiculer par les bateaux. On mène des recherches actives pour découvrir des surfaces répulsives à l'accrochage des balanes sur les coques de bateaux car leur présence en grand nombre augmente gravement la traînée.
La larve de Cirripède
Le schéma suivant représente leur cycle de vie. Au sortir de l'oeuf, la larve de Cirripède est une nauplius tout à fait ordinaire; au fur et à mesure de ses mues successives, elle se transforme en un petit organise enfermé dans sa carapace, la larve Cypris, mais ses antennes différencient une glande gluante. Cette larve nageuse se fixe alors à un support, et s'établit tête en avant et sens dessus-dessous. En plus de sa carapace normale, la Balane secrète autour de son corps des plaques calcifiées, aux angles saillants.
Ces plaques laissent entr'elles une ouverture, par laquelle l'animal peut étendre ses appendices plumeux, qui balaient l'eau comme un filet de pêcheur, pour prendre au piège les particules de nourriture et les petits animaux.
Céphalocarides, Branchiopodes, Ostracodes, Copépodes, Cirripèdes sont quelques-unes des Classes regroupant des petits Crustacés d'aspect primitif, mais en fait très spécialisés, et parfois de façon très spectaculaire. Cependant, les trois quarts des Crustacés forment la classe des Malacostracés, dont l'Ecrevisse fait partie et qui rassemble ceux qui nous sont les plus familiers.
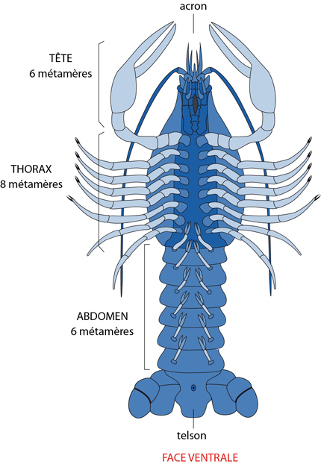
Les Malacostracés
En arrière de l'acron et des 6 métamères formant la tête, le tronc des Malacostracés est composé de 14 métamères et du telson; 8 d'entre eux forment un thorax souvent soudé à la tête. Tous les segments portent des appendices. Primitivement, les appendices thoraciques sont semblables: l'endopodite est la plus développée des deux branches, et sert à la marche et à la préhension. Puis, 1, 2 ou 3 paires de ces appendices thoraciques se tournent vers l'avant et se modifient en gnathopodes. Le thorax est couvert par une carapace, mais celle-ci est réduite dans certains Ordres de Malacostracés. Généralement, les branchies sont des épipodites modifiés.
Les appendices abdominaux antérieurs (souvent les 5 premières paires) sont semblables et biramés; ce sont des pléopodes, utilisés pour la nage ou le fouissage, pour faire transiter les particules alimentaires ou engendrer un courant d'eau respiratoire, pour retenir les oeufs pondus chez la femelle, ou parfois pour les échanges gazeux. Chez le mâle, une ou deux paires de pléopodes se transforment en organes copulateurs. En somme, leurs fonctions sont très variées.
Tout au bout, le dernier segment abdominal, le sixième, porte une paire d'appendices dont les 4 branches sont souvent étalées en palettes, entourant le telson, aplati lui aussi. Cette queue sert à la nage. Les variations autour de ce thème de base sont innombrables, d'où la systématique particulièrement ardue des Malacostracés.

Les Hexapodes
Le dernier groupe est aussi le plus diversifié. Il existe un nombre étonnant d'espèces d'Insectes: plus de 830.000 et peut-être 5, 10 ou 50 fois plus d'espèces à découvrir et à décrire encore. Ce groupe connaît un succès énorme, mais bien que l'on puisse citer divers caractères de leur organisation dont l'efficacité est évidente, il est délicat de mettre le doigt sur la caractéristique particulière à laquelle on puisse attribuer cette réussite remarquable. Leur plasticité évolutive entre sans doute en jeu: les Insectes proposent une variété de structures des plus étonnantes, qui montre autant d'adaptation à des milieux, des conditions de vie et des régimes alimentaires les plus variés. Les entomologistes distinguent deux douzaines d'Ordres actuels d'Insectes, plus une autre douzaine d'Ordres fossiles, à présent éteints. Nous n'entrerons pas dans les détails concernant tous ces Ordres
Les ailes sont bien entendu une caractéristique majeure des Insectes. Mais tous les Insectes ne sont pas ailés. Un groupe primitif aptère constitue les Aptérygotes qui regroupe notamment les Thysanoures et les Collemboles.

Le Thysanoure
Le Thysanoure représenté ici est le Lépisme ou « poisson d'argent », bien connu comme amateur de livres, mais c'est la colle de la reliure qu'il préfère.

Les Collemboles
Les Collemboles sont présents en nombres énormes dans les sols des prairies et des bois, parfois à la surface de l'eau, mais ils passent inaperçus, sauf des spécialistes qui les cherchent. Ils possèdent un appendice caudal bifurqué replié sous l'abdomen - la furca- dont l'extension soudaine catapulte le Collembole vers l'avant.

Les Ptérygotes
L’apparition des ailes chez les insectes est une innovation dont on découvre la première trace chez des fossiles qui datent d’il y a 325 millions d’années. Les insectes ailés constituent le groupe des Ptérygotes.
Le mécanisme évolutif qui a permis cette innovation n’est pas encore bien compris. Au cours de l'évolution, l’apparition des ailes s’est faite en plusieurs étapes. Les premiers fossiles Aptérygotes datent du Silurien. Les premiers Paléoptères sont apparus plus tard au Carbonifère, il y a –325 millions d’années. Delitzschala bitterfeldensis, découvert en 1996 en Allemagne, en est le plus ancien représentant connu. Les Libellules (ici en photo) et les Ephémères en sont des représentants actuels. Chez les Néoptères, plus récent, le mécanisme est perfectionné par l’ajout d’une articulation permettant la flexion des ailes le long du corps. C’est le cas par exemple chez la Locuste. Néanmoins, la question reste d’actualité car une aile fonctionnelle ne peut pas apparaître d’un coup par le résultat d’une seule mutation. Plusieurs hypothèses sont donc avancées. La première suppose qu’à l'origine, des petites expansions latérales du tergite, sur les segments thoraciques, auraient aidé l'Insecte à ne pas se retourner, ne pas tourbillonner quand il sautait. Puis ces petites plaques auraient grandi, lui permettant de sauter plus loin, en vol plané. Enfin, des articulations seraient apparues, assurant ainsi un vol battu, avec les ailes en mouvement. Une autre hypothèse donne comme fonction de départ de ces premières expansions un rôle de régulation thermique qui plus tard aurait évolué vers le vol. Les papillons actuels étalent encore largement leurs ailes au soleil afin que leur corps puisse atteindre plus rapidement une température permettant l’activité.
Puisque les ailes sont des évaginations du tégument, elles sont constituées de deux feuillets de cuticule. Le long d'une nervure, les deux cuticules sont épaissies, et séparées par une trachée; donc, on obtient une lumière tubulaire et de part et d'autre, deux renforcements cuticulaires; ainsi les nervures constituent un renfort squelettique efficace. Les nervures de l'aile s'ouvrent dans l'hémocoele, et sont emplies d'hémolymphe (c'est elle qui fait s'épanouir les ailes après la mue, en emplissant progressivement les nervures).
Si nous cherchons à retracer l'histoire évolutive des Insectes ailés, et par la même occasion à rendre compte de leur diversité actuelle, nous pouvons raisonnablement les classer en quatre groupes importants, allant des formes primitives aux plus évoluées. On se base sur des particularités de leur biologie, et aussi justement sur la structure des ailes, capables ou non de se replier sur l'abdomen et plus ou moins riches en nervures. Les Insectes Ptérygotes les plus primitifs possèdent des nervures disposées en réseau. Plus tard, on observera une tendance généralisée à la réduction de ce réseau en quelques grosses nervures longitudinales, reliées par des transversales.

Les Paléoptères
Les Paléoptères, rassemble des Ephéméroptères et les Odonates.
Les ailes elles ne sont capables que d'un mouvement vertical et ne peuvent être rabattues sur l'abdomen quand l'animal est au repos. Les Ephémères tirent leur nom de leur cycle de vie. Les larves vivent et croissent dans l'eau pendant des périodes qui atteignent 3 ans. Au terme de sa vie aquatique, la larve remonte à la surface, sa cuticule se fend sur le dos et libère un adulte ailé. La seule fonction de ce dernier est de s'accoupler au plus vite. Souvent, le laps de temps vécu dans le milieu aérien n'excède guère une journée. Les pièces buccales de l'adulte aérien sont atrophiées ; de toutes manières, il serait incapable de se nourrir.

Les Odonates
Les Odonates sont représentés aujourd'hui par les Libellules, comme celle-ci, et les Demoiselles. Ils ont une histoire fossile exceptionnellement longue et riche. La faune du Carbonifère contient notamment l'Insecte le plus gigantesque connu: une Libellule d'une envergure de 70 cm.

Les Polynéoptères
Le deuxième groupe est constitué des Polynéoptères. Néoptères comme les deux autres groupes qui suivront: c'est-à-dire que les ailes ont acquis une plus grande efficacité de mouvement car leur articulation est plus complexe. Au repos, les ailes peuvent se replier sur l'abdomen, et donc être en-dehors du chemin. C'est un événement important dans l'évolution de la Classe.

Hémimétaboles
Chez les Polynéoptères, pour lesquels les ailes sont renforcées par des nervures, c'est surtout la paire d'ailes postérieures qui semble avoir fait l'objet des plus grands soins: ce sont de belles structures en dentelle qui au repos peuvent être repliées en éventail ; dans de nombreux cas, la paire d'ailes antérieure ne représente guère que des organes durcis, protégeant la paire postérieure au repos. Les jeunes de ce groupe croissent en milieu terrestre plutôt que dans l'eau, mais on connaît peu de types larvaires spéciaux: le jeune fraîchement éclos n'est le plus souvent qu'une copie minuscule de ses parents, à l'exception des ailes qui apparaîtront plus tard. Au cours de la croissance, une série de mues assure l'évolution progressive vers la taille et l'organisation adulte. On dit qu’ils sont Hémimétaboles.

Les Polynéoptères
Les Ordres de Polynéoptères sont très variés, comme leurs modes d'existence; toutefois, leur régime alimentaire exige chez tous des pièces buccales de type broyeur, bâties sur le même plan général que celui du Criquet. Les criquets, les mantes religieuses, les phasmes, les blattes et les termites appartiennent à ce groupe.

Les Paranéoptères
Le troisième groupe d'Insectes est qualifié de Paranéoptères ; les ailes bien articulées présentent une certaine réduction du nombre des nervures. Mais chez certains, le vol perd de son importance. De nombreuses punaises ne volent qu'au moment de l'accouplement ou de la fuite. D'autres Paranéoptères comme les Poux, ont secondairement abandonné le vol. Comme chez les Polynéoptères, les jeunes éclosent sous une forme miniaturisée, mais essentiellement identique à celle de leurs peu engageants parents. On y retrouve entre autres les thrips, les pucerons, les punaises et les cigales.

Métamorphose
Le dernier groupe d'Insectes est le plus vaste par sa diversité. Un caractère qu'ils ont en commun, c'est l'abandon du développement direct, pour acquérir un cycle biologique plus complexe, doté d'un stade larvaire spécialisé, dont la niche écologique est généralement différente de celle de l'imago. On ne trouve plus ici de séries de mues avec passage progressif vers la taille et l'organisation de l'adulte. Au lieu de cela, l'individu mène successivement deux vies différentes. De l'œuf sort une larve, qui se nourrit, croît avec une série de mues, puis s'enferme dans une enveloppe, semble immobile et comme plongée en léthargie. En fait, à ce moment, l'anatomie tout entière de l'animal est remaniée intégralement, pour donner celle de l'adulte, qui s'échappe finalement de sa prison. C'est la métamorphose. Ce groupe est rassemblé sous la dénomination d'Oligonéoptères: le nombre de nervures principales de l'aile a encore diminué.
On y distingue 5 lignées majeures : les Lépidoptères, les Diptères, les Siphanoptères, les Hyménoptères et les Coléoptères. Chacune propose des spécialisations particulièrement remarquables. Ils représentent la majorité des espèces sur terre.
Les Arthropodes jouent un rôle considérable dans la vie des mers; sur terre et dans les airs, ils sont présents partout, innombrables tant en espèces qu'en individus. Il semble quelquefois que c'est nous, Vertébrés, qui vivons dans un monde d'Arthropodes où nous sommes tolérés.