Les platyhelminthes
Les 3 classes principales des Platyhelminthes montrent une adaptation remarquable tant à la vie libre qu'au parasitisme de plus en plus poussé. Nous étudierons la planaire, vers plat libre et aquatique, en situant précisément les dérivés cytologiques des 3 feuillets embryonnaires de ce premier Métazoaire triploblastique. Nous analyserons ensuite les caractéristiques de deux espèces parasites représentatives des deux autres groupes de platyhelminthes.
Enfin, nous ferons part des hypothèses à propos de l'origine et de l'évolution des Platyhelminthes.
Fiche récapitulative
- Métazoaires, triploblastiques, acoelomates ;
- À symétrie bilatérale ;
- Protostomiens ;
- Corps aplati, avec une partie antérieure (tête) enrichie en structures nerveuses et sensorielles ;
- Cavité gastro-vasculaire ouverte sur l’extérieur par un orifice (absente chez certaines formes parasites);
- Système d’excrétion différencié, les protonéphridies;
- Système nerveux formé d’une double chaîne ventrale, avec des parties plus développées (ganglions cérébraux) dans la partie antérieure;
- Reproduction sexuée et asexuée (fission, bourgeonnement);
- Hermaphrodites, avec des systèmes reproducteurs complexes;
- Marins, dulçaquicoles, ou terrestres;
- Libres ou parasites.
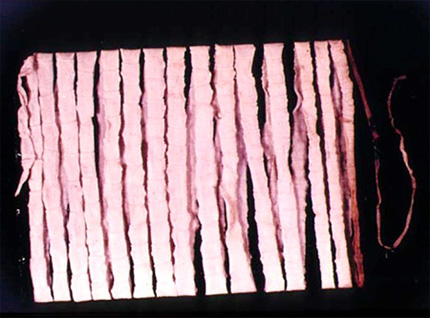
Ver solitaire
Comme leur nom le suggère, le groupe des platyhelminthes rassemble les vers au corps aplati dorso-ventralement. Ils sont généralement de petite taille, de 1 à 3 cm de long, bien que certaines formes parasites, comme les Taenia, puissent mesurer plusieurs mètres.\nCelui-ci a été extrait de l’intestin d’un homme, et enroulé sur une plaque.

Planaire marine
Les formes libres, comme les planaires, sont principalement aquatiques, habitant les eaux douces ou marines. Certains sont richement colorés et arborent des expansions de leur surface corporelle. Les formes libres se déplacent essentiellement sur leur surface ventrale, et peu sont capables de nager en pleine eau.

Les platyhelminthes terrestres
Les platyhelminthes terrestres sont rares, et confinés à des milieux très humides, comme ce ver coloré vivant dans les forêts tropicales humides.
2. Exemple(s)-type(s) - 2.1. La planaire, Dendrocoelum lacteum - 2.1.1. Examen externe
La planaire est un ver plat de 1 à 2 cm de long, de couleur blanchâtre. La faible épaisseur du corps permet de voir des structures internes par transparence.

Partie antérieure
La partie antérieure d'une planaire est élargie par deux petits lobes latéraux et porte ventralement un appareil adhésif (ventouse) et, dorsalement, deux taches pigmentées, les yeux.
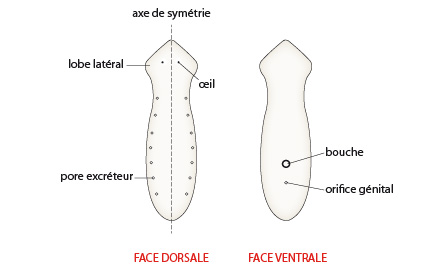
Planaire
La planaire présente une symétrie bilatérale de part et d’autre d’un axe longitudinal antéro-postérieur. Sur la face dorsale, on peut observer des paires d’orifices excréteurs.
La bouche n’est pas située dans la partie antérieure du corps, mais dans la partie médiane de la face ventrale du corps. Un peu plus en arrière, on observe un orifice génital communiquant avec les organes génitaux de la planaire.
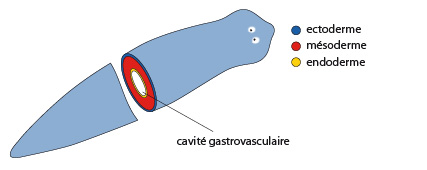
2.1.2. Examen interne
Cette planaire est découpée pour que nous y voyions les différents feuillets embryonnaires. Nous pouvons en observer trois. En plus de l'ectoderme, coloré ici en bleu, et de l'endoderme, coloré en jaune, présents chez les Cnidaires, un feuillet intermédiaire est inséré entre les deux précédents. C’est le mésoderme, coloré ici en rouge. A partir d’ici, nous conserverons ce code couleur pour indiquer l’origine mésoderme des tissus adultes. Au centre, la cavité gastro-vasculaire, entourée d’endoderme, est laissée blanche.
La coupe transversale nous démontre bien la structure triploblastique du corps. Les platyhelminthes sont acoelomates : on ne retrouve donc pas de cavité coelomique au sein du mésoderme.

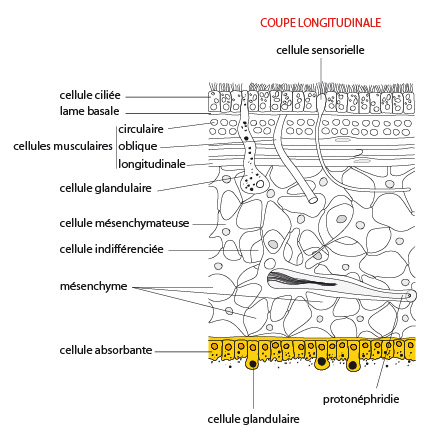
2.1.2.1. Tégument
Le tégument comporte un épiderme reposant sur une lame basale épaisse. L'épiderme est de type monostratifié et présente des cellules ciliées et des cellules glandulaires. Celles-ci déversent leur sécrétion muqueuse à la surface du corps qui est dépourvu de cuticule. Des cellules nerveuses sensorielles y sont également présentes. Sous l’épiderme, une lame basale épaisse confère une certaine rigidité au tégument.
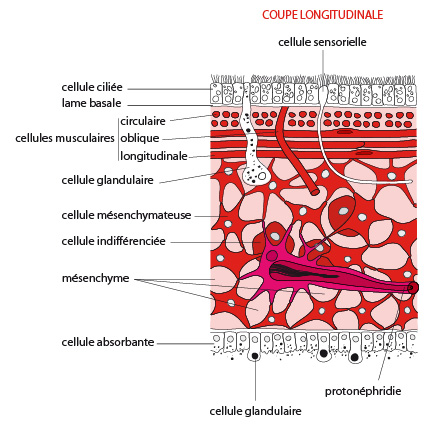
Mésenchyme
Sous cette lame basale, une couche musculeuse formée de cellules musculaires lisses agencées de manière assez peu organisée, dont l'orientation est principalement circulaire à l’extérieur et longitudinale à l’intérieur. Des cellules musculaires orientées obliquement joignent ces deux couches. Ces muscles permettent la déformation du corps.
L’espace compris entre les téguments et la cavité digestive est remplie d'un tissu constitué de cellules diverses, certaines libres, d’autres immobilisées, agencées en un mésenchyme présentant des espaces (lacunes) contenant du liquide interstitiel. Ce mésenchyme est issu du mésoderme, ce qui le différencie de la mésoglée des Cnidaires, cette dernière étant principalement d’origine ectodermique. La densité cellulaire y est nettement plus importante que dans la mésoglée des cnidaires, qui ne contenait que des cellules éparses dans une matrice fibreuse. Les cellules du mésenchyme sont impliquées dans le soutien et la mobilité du corps, et le stockage de réserves énergétiques. Certaines cellules de ce mésenchyme (les cellules indifférenciées) conservent la capacité de se différencier en un grand nombre de types cellulaires, et sont à l’origine de la grande capacité de régénération tissulaire chez ces vers. Nous verrons cela plus en détail dans la partie consacrée à la reproduction.
2.1.2.2. Système locomoteur
La planaire se déplace principalement par glissement à l’aide des cellules ciliées recouvrant sa surface ventrale. Le mucus déchargé par les cellules glandulaires sur cette même surface procure une lubrification facilitant le glissement de l’animal et un milieu visqueux contre lequel peuvent agir les cils. Elle utilise également les contractions de ses cellules musculaires pour modifier la forme de son corps et ainsi changer d’orientation. Cette musculature est également utile pour modifier l’épaisseur corporelle, lui permettant de s’insinuer dans des interstices du substrat.
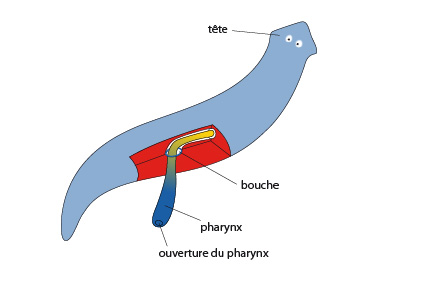
2.1.2.3. Système digestif
La planaire, comme de nombreux platyhelminthes libres, est carnivore. L’animal possède des récepteurs sensoriels capables de détecter des molécules dissoutes dans l’eau. C’est ce qui se produit lorsque le morceau de chair est déposé. Des molécules diffusent dans l’eau, la planaire les perçoit et se dirige vers leur source. Ensuite, un renflement apparaît à la surface ventrale du corps, à mi-chemin de l’axe antéro-postérieur de l’animal. Là se situe une bouche prolongée par un pharynx musculeux blanchâtre et évaginable. Cet organe est capable d’aspirer des proies. D’origine ectodermique, le pharynx est cilié et muqueux comme les téguments. Le pharynx mène au tube digestif, formé d'endoderme, qui est ramifié afin de distribuer les substances nutritives dans toutes les parties du corps. Celui-çi n'est ouvert que par l'extrémité buccale, et on ne parlera donc pas d'un véritable tube digestif, mais d'une cavité gastro-vasculaire, comme chez les Cnidaires.
La digestion
La digestion est initiée à l'extérieur du corps, par la libération d'enzymes protéolytiques, dans la cavité formée par le pharynx. La proie est ainsi partiellement digérée et réduite en un liquide avant son aspiration par les muscles pharyngiens.
La cavité gastro-vasculaire
La cavité gastro-vasculaire forme des extensions qui s’insinuent vers l’avant et l’arrière du corps, dans le mésenchyme. Sa paroi est formée de cellules glandulaires, qui déversent des enzymes digestifs, et de cellules absorbantes qui phagocytent les particules alimentaires et terminent le processus digestif grâce aux enzymes lysosomiales, après fusions des phagosomes avec des lysosomes.
Le matériel non digéré est éliminé par expulsion au niveau du pharynx. Les ramifications de la cavité gastro-vasculaire servent à accroître la surface digestive et absorbante.
2.1.2.4. Système respiratoire
Par sa forme aplatie et sa petite taille, la planaire dispose d’une grande surface corporelle par rapport au volume de ses tissus. En outre, la distance de diffusion des gaz respiratoires entre les cellules et le milieu extérieur demeure faible. Couplées à une activité métabolique qui reste limitée, ces caractéristiques permettent d’assurer un échange des gaz suffisamment rapide par rapport aux besoins de la planaire. Ainsi, la planaire ne dispose-t-elle d'aucun organe respiratoire spécialisé.
2.1.2.5. Système circulatoire
La cavité gastro-vasculaire s’insinue vers l’avant et l’arrière du corps, au sein du mésenchyme. Si cette complexité procure une extension de la surface intestinale propice à une digestion rapide, elle permet également la distribution des nutriments issus de la digestion dans l’ensemble du corps. Cette distribution est facilitée par les mouvements corporels. Il n’y a donc pas de système circulatoire différencié.
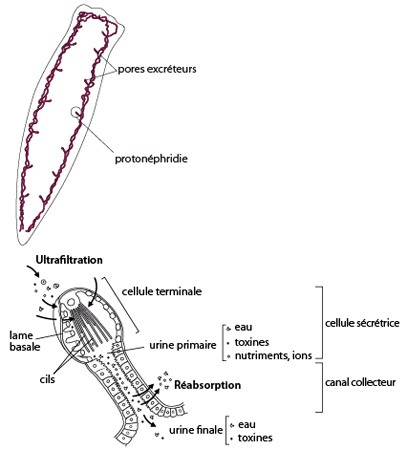
2.1.2.6. Système excréteur
Une des caractéristiques importantes des platyhelminthes est le développement de véritables organes excréteurs, les protonéphridies. Celles-ci sont formées de cellules bulbeuses, creusée d’une cavité, dites « à flamme » car à l'intérieur ondule un faisceau de cils évoquant une flamme. Par leur battement au sein de la cavité, les cils créent un flux d’eau qui transite depuis le mésenchyme vers l’intérieur de la protonéphridie au travers des cellules délimitant cette dernière. Disposées dans l’ensemble du mésenchyme, les protonéphridies déversent leur liquide dans un réseau de canaux collecteurs qui communiquent avec l’extérieur du corps par des pores, les néphridiopores. Notons que les déchets du métabolisme azoté, sont éliminés sous forme d'ammoniaque diffusant librement au travers du tégument, et non par les néphridiopores. Les protonéphridies ont essentiellement une fonction osmorégulatrice, visant à éliminer l’excédent d’eau douce qui pénètre en masse dans le corps de la planaire. Elles recueillent également les toxines en excédent dans les interstices des tissus.
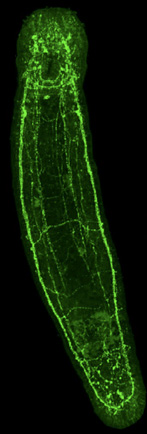
2.1.2.7. Système nerveux
Le système nerveux de la planaire a évolué avec la symétrie bilatérale et l’adoption d’une locomotion unidirectionnelle. Le système nerveux est constitué de deux cordons nerveux ventraux longitudinaux, reliés entre eux par des anastomoses, formant une sorte d’échelle alignée selon l’axe antéro-postérieur du corps. On les voit clairement sur cette image réalisée au microscope à fluorescence du turbellarié Macrostomum lignano, dans laquelle les cellules nerveuses sont mises en évidence grâce à des anticorps spécifiques de ces cellules.
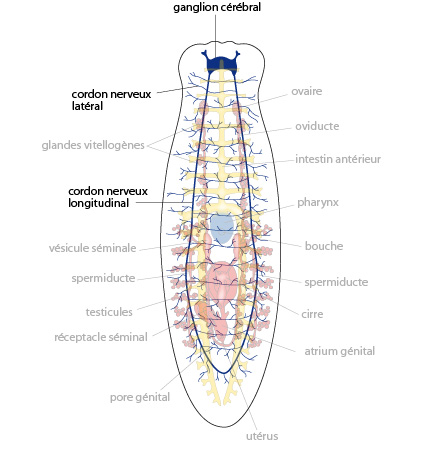
2.1.2.7. Système nerveux
De cette double chaîne nerveuse émanent des ramifications cellulaires qui irradient dans l’ensemble du corps.
La concentration en cellules nerveuses et sensorielles est plus grande dans la partie antérieure du corps, notamment dans les deux lobes latéraux de la tête. Dans la tête, les cellules nerveuses forment deux masses, les ganglions cérébraux, qui contiennent une quantité importante de corps cellulaires (péricaryons). Parmi les récepteurs sensoriels, on retrouve des cellules sensibles aux stimuli tactiles qui permettent à l’animal de percevoir la présence d’obstacles. Ils sont disposés sur l’ensemble du corps, avec une densité plus élevée au niveau de la tête et du pharynx. On trouve également des récepteurs aux substances chimiques présentes dans l’environnement (chémorécepteurs), particulièrement abondants sur les expansions latérales de la tête. Lorsqu’elle chasse, la planaire va faire osciller sa tête, balayant l’environnement chimique, de manière à percevoir d’éventuels gradients de substances qui lui permettraient de détecter des proies. Ce sont eux qui permettent à la planaire de s’orienter vers la source de nourriture.
Les monogonontes
La planaire possède également deux photorécepteurs assez complexes, appelés ocelles, placés sous l’épiderme, de part et d’autre de la ligne médiane de la tête. Ceux-ci sont constitués d’une cupule de cellules pigmentaires au sein de laquelle s’insinuent des cellules photoréceptrices dont la partie sensible est accolée de la couche pigmentée.
Cette disposition est proche de celle que l’on retrouve dans l’œil (dit inversé) des vertébrés. Les cellules photoréceptrices vont faire synapse avec des cellules nerveuses des ganglions cérébraux. De telles structures photoréceptrices sont incapables de former de véritables images. Leur rôle est plutôt de percevoir la direction d’une source lumineuse et de ses variations d’intensité. Selon l’origine du faisceau lumineux, la zone éclairée du fond de l’ocelle sera différente. Ce ne serait pas le cas si l’ocelle était une surface plane totalement accessible à la lumière. La planaire est un animal présentant un phototactisme négatif: elle va fuir les sources de lumière, préférant les endroits sombres où elle sera moins visible pour d’éventuels prédateurs.
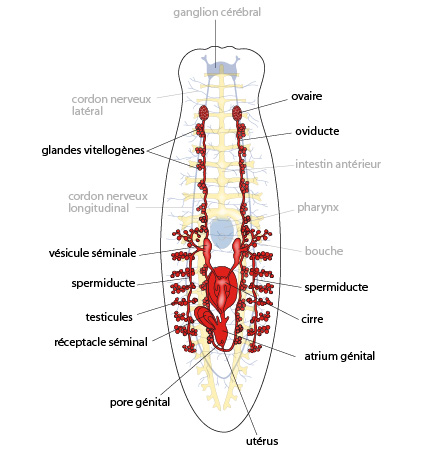
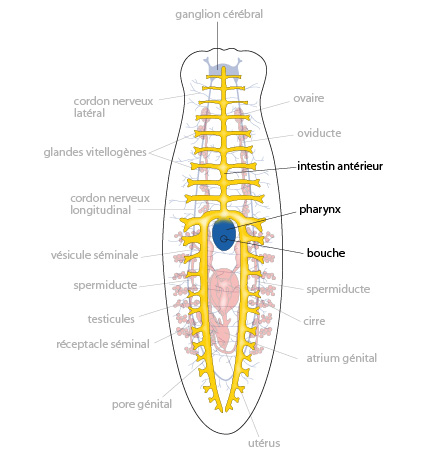
2.1.2.8. Système reproducteur
La planaire est hermaphrodite. Le système reproducteur, mâle et femelle, occupe un large volume. La partie mâle est constituée de nombreuses masses testiculaires dispersées au sein du mésenchyme. Ces testicules sont reliés à des tubules qui confluent en deux spermiductes qui se réunissent dans le plan médian, en une vésicule séminale, connectée au pénis (le cirre) logé dans une chambre musculeuse, l'atrium génital. Les organes génitaux femelles sont composés de dexu masses latérales, les ovaires, logés dans la partie antérieure du mésenchyme. Les cellules germinales qu’ils produisent transitent par deux tubes, les oviductes, dans lesquels des glandes vitellogènes déversent leur production de vitellus; les oviductes confluent en un utérus unique, connecté à l’atrium génital. Bien que ces animaux soient hermaphrodites, il y a accouplement, donc fécondation croisée, et non auto-fécondation.
Les spermatozoïdes sont déposés par le pénis d'un individu dans l'atrium génital de son partenaire; ils remontent les oviductes jusqu'aux ovaires, où se fait la fécondation.\nLes œufs fécondés descendent les oviductes, sont entourés au passage de cellules vitellines produites par les glandes vitellogènes, et débouchent dans l'atrium génital, où ils sont entourés d'une substance produite par l'utérus. Cette substance durcit et forme des cocons rigides, qui sont pondus par le pore génital.\nDans le cocon, les oeufs se développent et donnent en 2-3 semaines des petits vers semblables à leurs parents.
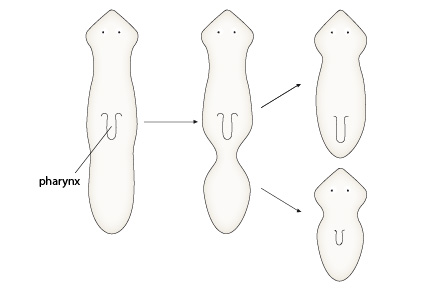
Les planaires
Les planaires peuvent aussi se reproduire de manière asexuée. Une constriction apparaît spontanément derrière le pharynx, et aboutit finalement à la séparation de deux individus complets. Ce phénomène est assez proche de celui de la régénération qui permet de restaurer une partie du corps ou d'un organe perdu accidentellement. La capacité de régénération de la planaire est exceptionnelle: en 1898, Morgan a montré qu’un 279ème d’une planaire était capable de régénérer un animal entier !
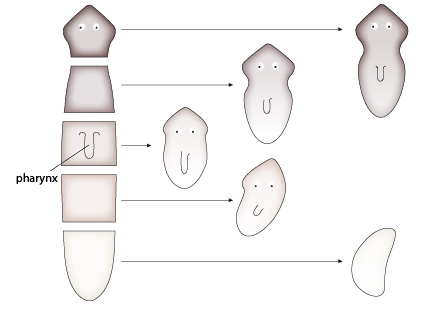
Les planaires
Procédons à une expérience. De multiples sections transversales sont réalisées au travers toute la longueur du corps d’une planaire.
1. Décrivez ensuite vos observations et tentez de répondre aux questions suivantes.
Question 1. Les segments conservent-ils la polarité originale ?
Oui, les segments conservent la polarité antéro-postérieure qu’ils avaient dans le corps de l’individu. En effet, pour chaque segment, le bord qui se trouvait du côté de la tête régénère une tête, tandis que le bord qui se trouvait du côté de la queue régénère une queue.
Question 2. La capacité de régénération est-elle la même de la tête à la queue ?
Non. Observez le fragment le plus antérieur. Celui-ci a régénéré une planaire complète, aux yeux et pharynx bien formés; en revanche, le morceau le plus postérieur n'a pu régénérer qu'une planaire informe, assez méconnaissable! Entre les deux extrémités, les planaires régénérées sont plus ou moins complètes: l'avant-dernière par exemple, n'a qu'un oeil... Cela indique que la capacité de régénération est plus grande du côté de la région antérieure, et diminue progressivement en direction de l'extrémité postérieure. Cela indique que les cellules de la partie antérieure du corps ont une pluripotence plus développée, qui peut aller jusqu’à la totipotence.
2. Dans une dernière expérience, des entailles longitudinales sont réalisées au niveau de la tête. Pensez-vous que l’individu qui résulte de cette régénération soit aussi efficace qu’avant cette opération ?
L’individu ainsi mutilé régénère souvent plusieurs têtes. Vivre avec plusieurs têtes ne doit pas constituer un avantage sélectif, bien au contraire! La capacité d’exploration du milieu et de réaction par rapport à un danger éventuel doit s’en trouver perturbée. En fait, les expériences montrent que la régénération d’une partie perdue accidentellement peut parfois s’avérer utile lorsque la régénération s’opère bien. Mais, elle est problématique dans la plupart des cas! En fait, les chercheurs pensent que la capacité de régénération va de pair avec la reproduction asexuée dont elle ne serait qu’un sous-produit. Le coût associé à la formation d’individus aberrants serait largement compensé par les bénéfices de la capacité de se multiplier par simple fission corporelle !
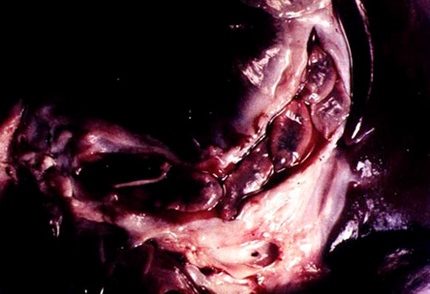
2.2. La douve hépatique, Fasciola hepatica
Comme on peut le voir dans la photo présentant une dissection du tube digestif d’un mouton infesté par des douves hépatiques, ce platyhelminthe parasite les canaux biliaires des Mammifères herbivores (mouton, vache,...). Elle peut parfois parasiter l'homme, s'il mange du cresson infecté.

2.2.1. Examen externe
La Douve mesure 2 à 3 cm de long. Comme la planaire, le corps blanchâtre de la douve est fortement aplati.

2.2.1. Examen externe
Sur ce schéma, on peut retrouver l’ensemble de ces structures. La bouche s'ouvre à l'extrémité antérieure du corps, au niveau du cône céphalique et est entourée d'une ventouse. Une seconde ventouse est présente ventralement, à proximité de l’orifice génital. A l'extrémité postérieure du corps et dorsalement, se trouve un pore excréteur. La minceur et la faible pigmentation des téguments laissent entrevoir les organes internes de la douve. L’orifice du canal de Laurer, situé dorsalement, reçoit l’organe copulateur mâle lors de l’accouplement.
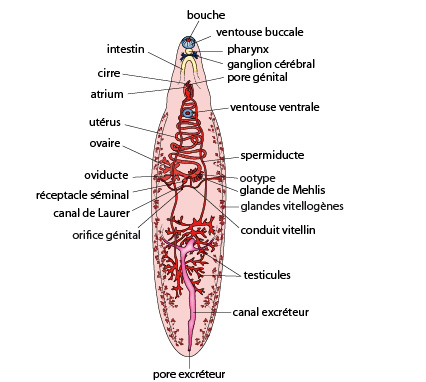
2.2.2. Examen interne
Organes internes d’une douve hépatique.
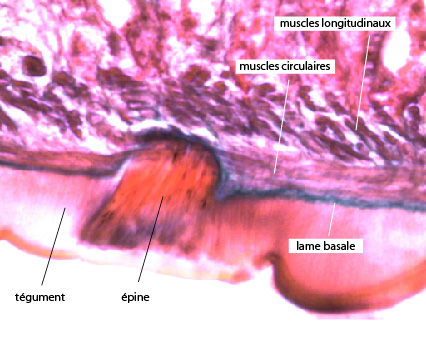
2.2.2.1. Tégument
Le tégument ne comporte pas un épiderme classique, comme chez la planaire. La couverture externe est formée d'une couche de cytoplasme syncytial riche en mitochondries et en vésicules de pinocytose. A sa surface, on remarque une absence totale de cils, et la présence de replis irréguliers qui augmentent la surface d’échange. Cette masse cytoplasmique est constituée d’expansions cytoplasmiques de cellules mésenchymateuses dont les corps cellulaires sont situés sous la lame basale, au-delà des couches de cellules musculaires. L’abondance de mitochondries et la présence de nombreuses vésicules de pinocytose dans ces expansions cytoplasmiques révèlent l’existence de transports actifs intenses, probablement de nutriments, prélevés chez l’hôte parasité. Il est probable que cette disposition particulière participe également à la protection de la douve contre les enzymes digestives et le système immunitaire de l’hôte. A la surface du corps, on observe des épines qui sont insérées dans le cytoplasme superficiel. Il est possible que ces structures servent à la libération de substances assurant une défense chimique vis-à-vis de l’hôte.
Mésenchyme
Le mésenchyme est semblable à celui de la planaire, avec ici et là des cellules musculaires lisses circulaires et longitudinales.
2.2.2.2. Système locomoteur
Le système locomoteur des douves est réduit par rapport à une planaire. L’absence totale de cils donne à la musculature un rôle prépondérant dans les déplacements qui demeurent néanmoins très réduits, et qui bénéficient des mouvements des fluides corporels de l’hôte. Une fois dans son hôte, la douve tend à bouger très peu, et s’arrime par ses deux ventouses aux tissus de l’hôte.
2.2.2.3. Système digestif
La douve se nourrit par deux voies. La première, associée à une pinocytose active par l’épithélium tégumentaire, a été décrite plus haut. La seconde consiste en un pompage des liquides corporels de l’hôte par le pharynx situé dans la partie antérieure. Le liquide ainsi absorbé va s’écouler via un court œsophage dans des diverticules de l’intestin, les caeca, dont la paroi est constituée de cellules absorbantes (les phagocytes) et de cellules glandulaires libérant des enzymes lytiques. La digestion est donc en partie extracellulaire.
2.2.2.4. Système respiratoire
La douve ne possède aucun système respiratoire propre. Les gaz respiratoires sont échangés à tout endroit de la surface corporelle. Notez que la douve est capable de survivre avec des quantités d’oxygène extrêmement faibles, voire nulles, comme cela peut se produire localement dans les voies digestives de l'hôte. Dans ces circonstances, elle dépend d’une alimentation énergétique basée sur la glycolyse anaérobie.
2.2.2.5. Système circulatoire
Comme mentionné dans la description du système digestif, la cavité gastro-asculaire assure la fonction de système circulatoire pour les nutriments. Il n’y a pas d’organe spécifique du système circulatoire.
2.2.2.6. Système excréteur
Comme la planaire, la douve possède des protonéphridies, connectées à des tubules confluant vers un pore excréteur unique. Elles interviendraient dans l’excrétion de l’ammoniaque et l’osmorégulation.
2.2.2.7. Système nerveux
Le système nerveux de la douve est très proche de celui de la planaire, avec une double chaîne nerveuse ventrale anastomosée. De nombreux récepteurs tactiles sont concentrés autours des ventouses. En revanche, la douve ne possède pas d’ocelles.
2.2.2.8. Système reproducteur
Comme la planaire, la douve est hermaphrodite. Le système reproducteur mâle et femelle occupe la majeure partie du corps. Deux masses testiculaires ramifiées sont placées dans la partie postérieure de l’individu et connectées par deux spermiductes qui se déversent dans une vésicule séminale commune, associée à un appareil copulatoire évaginable, le cirre. Le système femelle comporte un seul ovaire, connecté à un oviducte, aboutissant dans l’ootype pour repartir dans l’utérus contourné. De très nombreuses glandes vitellogènes sont dispersées autours des masses testiculaires. Elles déversent leurs productions, du vitellus, au niveau de l’ootype, via deux canaux vitellins. Les glandes de Mehlis, entourant l’ootype, y sécréteraient un mucus utile à la lubrification de l’utérus. Le système reproducteur mâle et femelle aboutit tout deux au niveau d’un atrium génital.
Lors de l’accouplement, chaque partenaire introduit son cirre dans le pore génital ventral de l'autre partenaire. Le sperme est amené dans un réceptacle séminal, connecté à l’oviducte, et les deux partenaires se séparent. Ensuite, les ovocytes passent dans l’oviducte et seront fertilisés par le sperme venant du réceptacle séminal au niveau de l’ootype, puis recouverts de cellules vitellines, avant d’être recouvert d’une enveloppe et transférés dans l’utérus. Dans certains cas la douve peut insérer son cirre au niveau de l’orifice dorsal du canal de Laurer aboutissant au réceptacle séminal. Une douve peut produire des quantités énormes d’œufs, jusqu’à plus de 100,000 fois la quantité produite par une planaire !
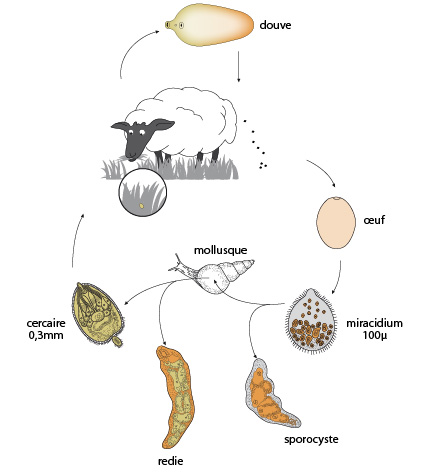
Cycle de vie de la douve
La douve adulte, qui mesure quelques centimètres, vit dans les canaux biliaires d’un mammifère, dont l’homme.
Lorsqu’elles sont nombreuses, elles peuvent causer de sérieux problèmes... Les œufs qu’elles produisent vont être envoyés dans les voies digestives de l’hôte, et passer dans les excréments de ce dernier. S’il atterrit dans l’eau, l’œuf éclot et forme une larve, le miracidium (ou larve miracidie).
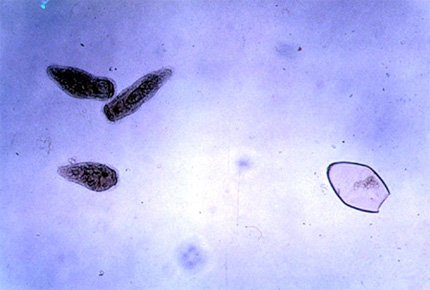
Larves miracidies
Sur cette photo, on peut voir à droite, un œuf éclos, et trois larves miracidies à gauche. Cette larve, ciliée et nageuse et longue d’une centaine de micromètres, présente une ébauche de système digestif, un système nerveux, des yeux, des muscles, et des amas de cellules indifférenciées dans la partie postérieure. La larve doit rapidement trouver un hôte, un mollusque de type limnée, et pénétrer dans son poumon pour continuer son développement. Là, elle se transforme en un autre stade de développement, le sporocyste. Ce stade est très simplifié: pas d’yeux, de cils, ni de tube digestif, mais un grand nombre de cellules germinales qui, par multiplication, donnent naissance à des larves rédies.
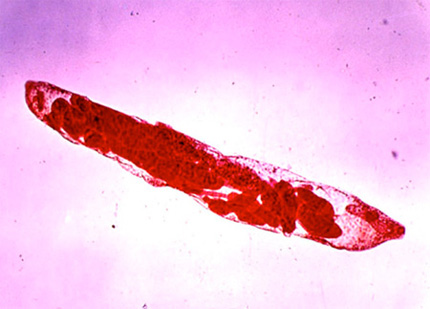
Larves rédies
Ces larves rédies, pourvues cette fois d'un système digestif simple, et contenant encore des cellules embryonnaires, crèvent le sporocyste, et migrent depuis le poumon vers la glande digestive de leur hôte mollusque, où elles se nourrissent, grossissent, bourgeonnent pour former de nouvelles générations de rédies. Puis après un certain nombre de générations, les rédies forment en elles des cercaires. Ces douves miniatures de 300 µm, possèdent une bouche, un système digestif et un appendice caudal, leur procurant une certaine mobilité.

Larves cercaires
Ces larves cercaires sortent du mollusque en perforant ses tissus et nagent vers une plante aquatique, perdent leur queue, et sécrètent une enveloppe kystique. Ce stade enkysté est appelé métacercaire.
Cycle de vie de la douve
Ce stade ne continuera son développement que lorsque la métacercaire sera avalée par un hôte convenable; à ce moment, la paroi du kyste est perdue, et la jeune douve se développe dans le tube digestif de l’hôte définitif où elle gagne les canaux biliaires et devient adulte.
Chaque œuf pondu par la Douve adulte pourrait donner naissance à des dizaines de milliers de cercaires, étant donné la multiplication asexuée qui se produit dans le sporocyste et la rédie. Or la Douve pond des milliers d'œufs par jour ! L’hôte intermédiaire (mollusque) et les hôtes définitifs (mouton, vache, ... homme) se retrouvant dans le monde entier, la douve est cosmopolite.
Les Douves irritent le foie, les canaux biliaires et produisent une atrophie hépatique. Le bétail en est très affaibli et peut même en mourir. Chez l'homme, cette parasitose, appelée distomatose, est redoutable par sa durée. Elle occasionne une fatigue, des douleurs abdominales et une fièvre continue.

3. Origine, diversité et évolution
Quatre classes de platyhelminthes ont été identifiées. Les turbellariés, les trématodes, les monogènes et les cestodes.
La planaire est représentative des turbellariés, qui comprennent principalement des formes aquatiques libres. La douve hépatique appartient elle à la classe des trématodes, dont la plupart sont des endoparasites (qui vivent à l’intérieur de l’hôte). Les monogènes sont des vers principalement parasites externes (ectoparasites) recouverts d’un tégument semblable à celui de la douve. Enfin, les cestodes, dont le taenia est un des représentants les plus connus, forment une quatrième classe de vers plats exclusivement endoparasites.
Décrivons rapidement la structure d’un cestode parasite de l’homme, le ver solitaire ou taenia. Ce ver parasite l'intestin grêle de l'Homme à l'état adulte, et les muscles du Cochon, à l'état larvaire. Il mesure 2-3 mètres de long mais n'atteint au maximum qu'un cm de large. La partie antérieure est un petit renflement en massue, le scolex, gros comme une tête d'épingle et muni au sommet d'une couronne de crochets rigides et de 4 ventouses latérales.
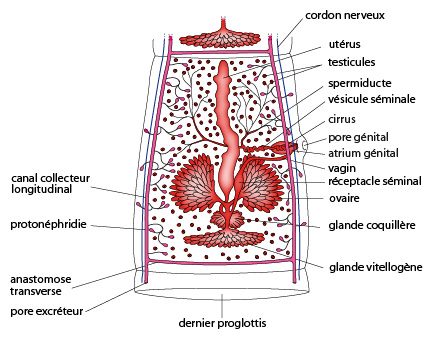
Les proglottis
Le reste du corps est composé d'environ un millier de segments identiques, les proglottis, provenant d'un bourgeonnement continu du scolex. Le tégument, semblable à celui de la douve, protège à la fois le ver des sucs digestifs de son hôte, et permet l'absorption des nutriments.
Le système nerveux comporte 2 ganglions cérébraux anastomosés dans le scolex, prolongés par deux cordons nerveux latéraux qui font toute la longueur du corps. Ce dernier ne présente aucune cavité gastrovasculaire.
Un appareil excréteur formé de protonéphridies associées à des canaux sont présentes dans chaque proglottis et aboutissent à l’extrémité postérieure de l’individu.
Le système génital hermaphrodite est construit selon un schéma semblable à celui de la douve. Les organes des deux sexes sont bien développés dans les segments moyens; en revanche, les proglottis antérieurs, plus jeunes, sont plutôt mâles. L'accouplement se réalise entre des proglottis antérieurs fonctionnellement mâles et des moyens, fonctionnellement femelles.

Les proglottis
Par cette disposition, les proglottis les plus postérieurs contiendront uniquement un utérus remplis d'oeufs, nommés les proglottis cucurbitains. Ces cucurbitains peuvent se détacher, et transiter avec le bol alimentaire vers l’anus. Une fois éliminés dans les fèces, ces proglottis peuvent se désintégrer et libérer des milliers oeufs, dont le développement peut démarrer s’ils sont ingérés par un nouvel hôte.
Exercice
Avant de poursuivre, faites l’exercice suivant : recherchez, chez le taenia, les caractéristiques communes avec la planaire et la douve..
Origine et évolution
L’origine et l’évolution au sein du groupe des platyhelminthes sont sujettes à d’intenses débats au sein de la communauté des biologistes. L’existence même d’un clade qui rassemblerait tous les vers classés parmi les platyhelminthes est sujette à caution. En effet, aucune synapomorphie autre que la forme aplatie du corps ne permettrait de les distinguer des autres animaux triploblastiques! Néanmoins, on peut également ajouter comme caractère dérivé propre que ce sont les seuls organismes triploblastiques ayant un système digestif à un seul orifice.
Les platyhelminthes sont des animaux à symétrie bilatérale, appelés Bilateria. Cette symétrie était déjà présente chez certains cœlentérés dérivés, comme les anthozoaires (au sein des Cnidaires). Les platyhelminthes présentent les synapomorphies propres aux animaux triploblastiques: la présence du mésoderme, la céphalisation et la centralisation du système nerveux, et la présence d’organes vrais.
Cependant, la simplicité morphologique de certains platyhelminthes, et la difficulté de mettre en évidence des synapomorphies propres à tous les platyhelminthes soutient l’idée que leur ancêtre serait à l’origine, non seulement de tous les platyhelminthes actuels, mais aussi de tous les protostomiens actuels. Dans ce cas, le phylum des platyhelminthes serait un groupe paraphylétique, puisque leur phylum ne comprendrait pas toutes les lignées de leur ancêtre.
Quelle pouvait être la structure de cet ancêtre ?
Actuellement, deux théories s’affrontent pour expliquer les origines des triploblastiques.
Selon la première théorie, l’absence de cavité corporelle pourrait être un caractère primitif (plésiomorphie) présent chez les premiers animaux triploblastiques. Cet ancêtre aurait donné naissance aux platyhelminthes modernes, ainsi qu’à tous les autres organismes triploblastiques dont certains auraient acquis une cavité corporelle coelomique.
Une autre théorie suggère que les premiers organismes triploblastiques possédaient une cavité corporelle, le cœlome, creusée au sein du mésoderme. Nous en reparlerons dans les chapitres suivants. Les platyhelminthes pourraient avoir perdu cette cavité et adopté une forme aplatie. Nous verrons par la suite qu’une manière de perdre ce coelome serait de conserver des caractéristiques propres à la larve, dans un processus qualifié de néoténie. Dans ce cas, l’absence de cavité serait une synapomorphie des platyhelminthes qui ne serait plus paraphylétique.
Questions
1. De nombreux organismes d’eau douce limitent les problèmes d’entrée d’eau par une réduction de la perméabilité des téguments. Pourquoi les platyhelminthes n’ont-ils pas recouru à cette stratégie ?
2. Les quelques espèces de platyheminthes terrestres sont confinés à des milieux extrêmement humides. Quelles en seraient les raisons ?
3. En comparant la planaire et la douve, identifiez les adaptations de la douve au parasitisme.
4. Identifiez les avantages sélectifs, et les éventuels inconvénients, du cycle vital de la douve hépatique.
5. Chez les cestodes de petite taille, la fécondation croisée est la règle. Pas chez les espèces de grande taille. Comment expliqueriez-vous cette différence ?