Les Mollusques
La diversité des Mollusques est étonnante; ils se sont adaptés aux habitats les plus variés. A première vue, il est bien difficile de leur trouver à tous des caractères communs. Pourtant, une comparaison attentive permet de tracer le schéma d'un "ancêtre hypothétique". C'est par là que nous commencerons.
Nous choisirons ensuite un exemple concret dans les Classes principales : Gastéropodes, Lamellibranches, Céphalopodes et Scaphopodes. L'examen de leur structure (examen externe et systèmes digestif, locomoteur, respiratoire) vous mènera à critiquer le modèle ancestral proposé.
Enfin, nous poserons la question de l'origine et l’évolution du groupe des Mollusques.
Fiche récapitulative
- Métazoaires triploblastiques coelomates ;
- Cavité corporelle de type hémocoele, avec une réduction importante du coelome ;
- À symétrie bilatérale primaire, pouvant tendre secondairement vers l’asymétrie ;
- Métamérie inexistante ou réduite ;
- Adulte présentant 3 zones superposées : un pied ventral musculeux, une masse viscérale dorsale, un manteau sécrétant une coquille (interne ou externe) ou des spicules calcaires ;
- Système digestif complet avec régions spécialisées, présence d’une paire de cordons nerveux ventraux, système circulatoire ouvert (sauf les céphalopodes) avec présence d’un cœur, système excréteur complexe (néphridies) ;
- Reproduction sexuée (le plus souvent hermaphrodisme et fertilisation croisée) ;
- Larve trochophore + larve véligère ;
- Clivage spiral holoblastique ;
- Terrestres, marins ou dulçaquicoles.

Présentation du groupe
Le groupe des mollusques comprend une grande diversité de forme: un escargot, une moule et un poulpe sont trois types de mollusques bien différents... On peut néanmoins les caractériser comme des animaux à corps mou, généralement protégés par une coquille interne ou externe.

Les nucelles
La plupart des formes actuelles sont marines, tandis que les espèces d’eau douce sont peu nombreuses. Sur cette photo vous voyez des nucelles que vous verrez au stage de biologie marine. Ce mollusque gastéropode mange d'autres mollusques et des balanes dans les parties moyenne et basse de l'estran.

Les gastéropodes
Seuls les gastéropodes, groupe auquel appartiennent les escargots et les limaces, ont colonisé la terre ferme.
2. Le mollusque ancestral hypothétique - 2.1. Examen externe
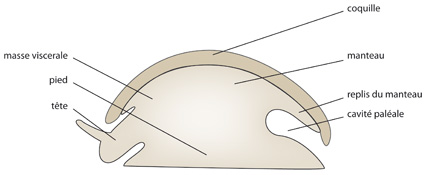
A première vue, les Mollusques semblent très différents les uns des autres. Pourtant, ils sont tous construits sur le même plan de base. On connaît peu de chose sur l'organisation anatomique des premiers mollusques à partir desquels se sont diversifiées toutes les formes actuelles. Pour essayer de bien comprendre ce qu'est un Mollusque et ensuite leur diversification, nous commencerons par examiner un Mollusque ancestral hypothétique reconstitué par inférence en utilisant nos connaissances actuelles. Sa structure fondamentale pourrait se schématiser comme sur ce dessin.
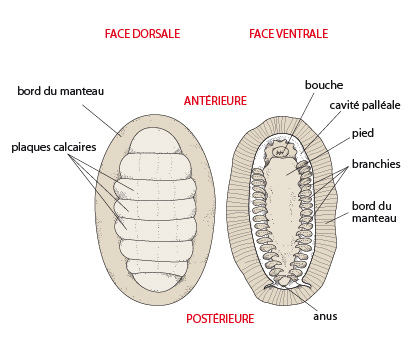
Dans le corps du Mollusque, on reconnaît trois régions : la tête, le pied, et la masse viscérale ; le tout est recouvert par le manteau, qui sécrète une coquille. Le manteau forme un repli sur tout le pourtour du corps. Attention ! Ce que vous regardez est une coupe. Imaginez ce schéma en trois dimensions. Le repli du manteau ménage un espace libre entre le corps et lui-même : c'est la CAVITE PALLEALE.
Le Mollusque ancestral devait présenter une symétrie bilatérale. Selon toute probabilité, le Mollusque ancestral était herbivore et se nourrissait d'algues ; il possédait un appareil digestif ressemblant à celui que l'on trouve chez la majorité des Mollusques actuels.
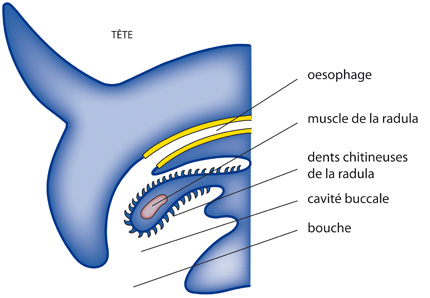
La radula
La bouche s'ouvrait dans une cavité buccale dont le plancher était occupé par une masse musculaire (l'odontophore) portant une structure typique des Mollusque : la radula. Ce ruban membraneux est muni de dents chitineuses réparties en rangées parallèles.

Nutrition de la radula
Des muscles rétracteurs et protracteurs associés à la radula et au tissu cartilagineux qui la porte, permettaient de projeter la radula et de la mouvoir contre les aliments, à la manière d’une rape. Ceci permet entre autre de brouter les algues sur les rochers.

Radula de patelle
La photo du haut présente la vue ventrale d'une radula de patelle photographiée au microscope. Notez la disposition régulière et l'orientation des dents. Observez également la patelle sur le rocher, photographiée lors de la marée basse en stage de biologie marine.

2.2. Examen interne - 2.2.1. Système digestif
Voici le système digestif du supposé mollusque ancestral. Commençons par la partie antérieure. Une paire de glandes salivaires s'ouvre dans la cavité buccale. Ces glandes sécrètent notamment du mucus qui lubrifie la radula et englobe les particules alimentaires.
La nourriture passe ensuite de la cavité buccale dans un oesophage tubuleux qui s'ouvre dans un estomac conique. Les petites particules alimentaires empruntent les canaux des glandes digestives associées à l'estomac pour y subir une digestion intracellulaire. Chez la plupart des espèces actuelles il s'opère une certaine digestion extracellulaire par des enzymes déversées dans la cavité stomacale, mais ce perfectionnement n'existait probablement pas chez l'ancêtre. L'intestin fait suite à l'estomac. L'anus s'ouvre postérieurement dans une cavité, la cavité palléale.
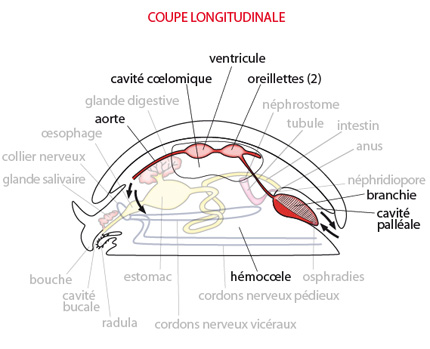
2.2.2. Système respiratoire - 2.2.3. Système circulatoire
Dans le système circulatoire et respiratoire, le coeur consiste en une paire d'oreillettes postérieures et un ventricule simple antérieur. Il est placé dans la cavité coelomique localisée dans la partie dorsale du corps. Notez que le coelome entoure le coeur ainsi qu'une portion de l'intestin ; c'est donc un coelome péricardique et périviscéral.
Les système circulatoire et respiratoire
Dans un tel coeur, les oreillettes drainent le sang venant des organes respiratoires (les branchies ou cténidies) et ce sang oxygéné passe ensuite dans un unique ventricule musculeux, qui l'envoie dans une artère dorsale, l’aorte. Cette dernière se prolonge par des vaisseaux sanguins plus petits qui distribuent le sang dans des lacunes baignant immédiatement les tissus ; il s'agit donc d'un système circulatoire ouvert : les lacunes n'ont pas de parois propres, ce sont des espaces au contact direct du liquide extracellulaire des tissus. Cette cavité générale du corps est appelée l’hémocoele.
Comme chez les mollusques actuels, le sang contenait un pigment respiratoire structurellement proche de l'hémoglobine, à la différence que son hème contient du cuivre au lieu de fer, ce qui lui donne une couleur bleue. Ce pigment est appelé HEMOCYANINE en référence à sa couleur bleue cyan.
Les branchies du Mollusque ancestral hypothétique sont localisées dans la cavité palléale et sont également appelées cténidies. Elles devaient être formées d'un axe central et de filaments s'attachant de part et d'autre de cet axe, ressemblant à des « peignes ». La position de ces cténidies divise la cavité palléale en une chambre supérieure et une chambre inférieure. Par le courant inhalant, l'eau entrant dans la chambre inférieure, passe à travers la cténidie, arrive dans la chambre supérieure et ressort par le courant exhalant. Le courant d'eau est engendré par les cils qui garnissent la surface des filaments.
Le sang des mollusques est oxygéné au niveau de ces cténidies. Deux vaisseaux sanguins traversent l'axe de la cténidie : le vaisseau sanguin afférent ramène le sang venant du corps, qui parcoure alors les filaments, puis emprunte le vaisseau sanguin efférent. Le sang oxygéné passe ensuite dans les oreillettes du coeur, puis le ventricule, puis l'aorte pour se déverser dans l’hémocoele.
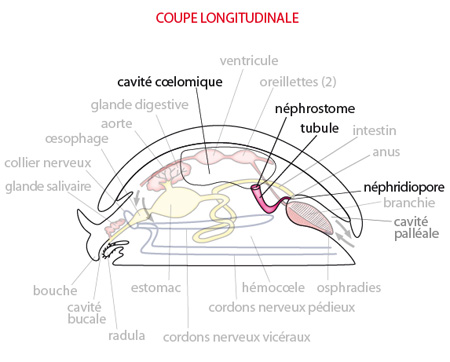
2.2.4. Système excréteur
Les organes excréteurs du Mollusque ancestral consistaient probablement en une paire de reins appelés néphridies.
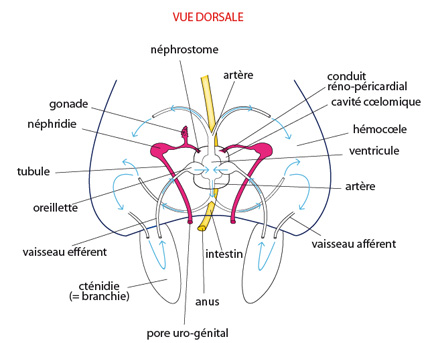
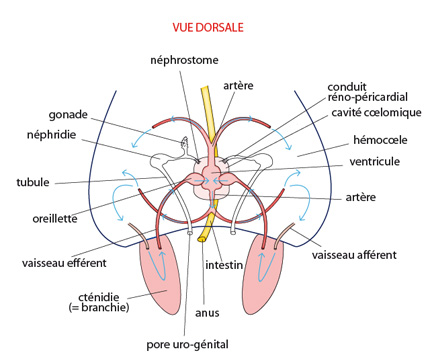
Les système excréteur
Une des extrémités de chaque néphridie est ouverte dans le coelome par un néphrostome; l'autre extrémité s'ouvre à l'extérieur, dans la cavité palléale, par le néphridiopore. Les deux néphridiopores gauche et droit s’ouvrent de part et d'autre de l'anus.
Le coelome (ou cavité coelomique) reçoit des déchets de deux sources: le coeur y libère un filtrat sanguin et certaines glandes du péricarde y sécrètent des déchets. Le liquide passe alors dans la néphridie via le néphrostome. Une certaine réabsorption de substances utiles vers l’hémocoele a lieu au niveau du tubule, avant que l'urine finale ne soit déversée dans la cavité palléale par le néphridiopore.
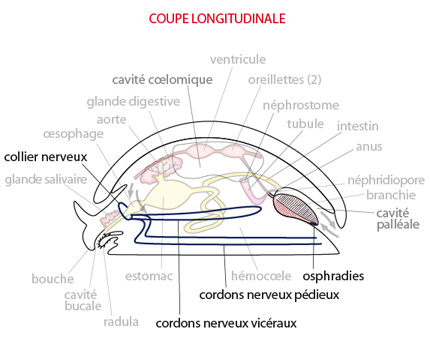
2.2.5. Système nerveux
Le système nerveux du Mollusque primitif se compose d'un collier nerveux entourant l'oesophage; deux paires de cordons nerveux s'en échappent et se dirigent vers l'arrière du corps. La paire ventrale innerve les muscles du pied: ce sont les CORDONS PEDIEUX. La paire plus dorsale innerve le manteau et les organes viscéraux : ce sont les CORDONS VISCERAUX.
Les mollusques sont donc bien des tétraneures.
La perception de l'état de l'environnement se fait au travers d'une série d'organes des sens, pas toujours présents chez toutes les espèces actuelles: des cellules olfactives, des yeux, des tentacules, des statocystes (organes de l'équilibre) et des osphradies. Ces dernières sont des chémorécepteurs localisés à la partie postérieure des branchies, et sensibles à la qualité et la turbidité de l'eau qui arrive avec le courant inhalant.
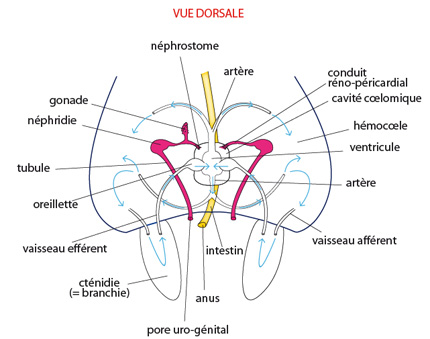
2.2.6. Système reproducteur
Dans leur organisation essentielle, les systèmes reproducteurs des Mollusques actuels sont proches de celui du Mollusque ancestral.
Chez le Mollusque ancestral, une gonade s'étend dans la partie dorso-latérale antérieure. A maturité, les ovocytes ou les spermatozoïdes sont déversés dans la cavité coelomique, et transportés vers l'extérieur par les canaux des néphridies. Le néphridiopore est d’ailleurs également appelé pore urogénital. La fécondation est externe; elle a lieu dans l'eau de mer environnante.

Les groupes de Mollusques
Maintenant, nous allons étudier plus en détail les principaux groupes de Mollusques actuels. Nous nous limiterons à une structure de base, dans un premier temps. En cours de route, faites continuellement le rapprochement avec leur ancêtre hypothétique, que nous venons d'examiner.

3. Exemples types
Ce schéma illustre clairement la distinction entre différents groupes de mollusques que nous allons voir par la suite. Les deux premiers dessins représentent respectivement les polyplacophores et monoplacophores. Ces mollusques ont la tête, le pied et la cavité palléale complètement recouverts par le manteau et la coquille. Puis, on retrouve les gastéropodes et les céphalopodes. Ces 2 groupes ont une tête distincte et très développée, un manteau qui n’entoure que la partie viscérale et leur tube digestif en forme de U. Les derniers schémas représentent respectivement les bivalves et les scaphopodes qui ont une tête régressée ou absente et le manteau quant à lui, entoure la totalité de l'animal.

4. Exemple type 1 : les Polyplacophores (et monoplacophores)
La Classe des Polyplacophores contient les Chitons. Ce sont des animaux aplatis qui vivent à faible profondeur en zone littorale intertidale et que l’on retrouve surtout sur les substrats durs.

Les chitons
Les chitons sont solidement fixés au substrat par leur pied. Il est nécessaire d’utiliser un outil (ici couteau) afin de pouvoir prélever l’organisme. Observez sur cette photo la taille relativement réduite (entre 2 et 4 cm) des chitons trouvés sur nos côtes.
Les chitons
Leur particularité est la constitution de la coquille en 8 plaques calcaires articulées entre elles. Le bord des plaques est souvent recouvert par le manteau. Dans ces plaques dorsales on retrouve des organes sensoriels photorécepteurs nommés les esthètes qui renseignent l’animal sur la quantité de lumière. Les chitons n’ont pas d’yeux céphaliques. Le pied large et ovale sert de ventouse à l'animal pour se fixer aux rochers et pour s'y déplacer lentement, en râpant les algues par des coups répétés de sa radula constituée de plusieurs rangées de 17 dents. Le bord du manteau s'étale tout autour du pied et est bordé de petites plaquettes calcaires. Lorsque les rochers sont découverts, les chitons s’adhèrent fermement au substrat et les plaques recouvrent complètement leur corps pour éviter le dessèchement. Leurs plaques articulées leur permettent également de s’enrouler sur eux-mêmes comme les cloportes quand ils se détachent de leur support. On pourrait penser que les plaques de la coquille sont l'indice d'une métamérisation, mais ces plaques ne correspondent ni à la disposition des multiples paires (6 à 88 selon les espèces) de branchies buissonnantes disposées le long du pied, sous le manteau, ni aux commissures transversaux entre les cordons nerveux longitudinaux. Aucun des autres organes internes ne montre de disposition suggérant une segmentation. Vous risquez d’ailleurs de voir des chitons lors du stage de biologie marine, soyez attentif. L'intérêt majeur du groupe des polyplacophores est qu’il présente une allure segmentée, dont il faut se méfier lors de l'interprétation tout comme les monoplacophores.
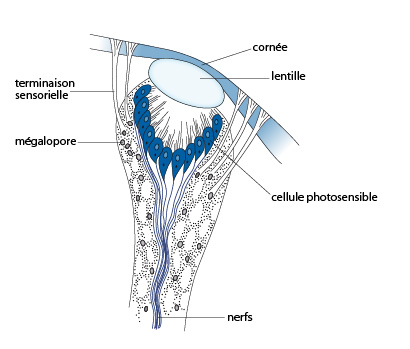
Les esthètes
Ces esthètes, un caractère propre à ce groupe des polyplacophores, se trouvent en grand nombre dans les plaques dorsales du polyplacophore et renseignent l’animal sur la quantité de lumière. Ce sont des cellules du manteau qui montent le long de canaux verticaux et se développent dans des pores (mégalopores) s’ouvrant dans les dernières couches de la coquille.
Les monoplacophores
Un groupe proche des polyplacophores est celui des monoplacophores, un groupe qui ne contient que très peu d’espèces vivantes et ayant une coquille patelloïde. Ils vivent sur les fonds océaniques et démontrent une anatomie interne sériée que vous verrez en fin de ce chapitre.

5. Exemples-type 2 : les Gastéropodes - 5.1. Examen externe
Ici nous présentons le groupe des gastéropodes qui est fort bien connu. Au sein des gastéropodes on fait une distinction entre les prosobranches, les gastéropodes qui ont des branchies en avant du corps, les opisthobranches avec leurs branchies à l’arrière du corps et les pulmonés, étant des gastéropodes terrestres qui ont acquis une sorte de poumon, le pneumostome.
Vous voyez ici une Limace, gastéropode terrestre. Son nom est Arion rufus, ou Arion roux. Sa couleur varie de l'orangé pâle au noir, en passant par le rouge brique et le brun. C'est une de nos plus grandes Limaces: elle peut atteindre 15 centimètres de longueur. On en trouve dans les bois, les champs, les prés et les jardins, où elle dévore des légumes, des fruits, des céréales et des champignons. Faites attention, le terme limace désigne en français les gastéropodes sans coquille externe.
La limace, adaptée à la vie terrestre, craint malgré tout la sécheresse; par temps sec, elle est nocturne; par temps humide, on peut la rencontrer en plein jour. On en conclut qu'elle recherche l'humidité et la fraicheur, mais qu'elle ne craint pas spécialement la lumière. Sa vie dure en moyenne un an. Les jeunes éclosent en automne puis s'enfoncent dans l'humus pour se protéger du gel. Ils remontent au printemps et atteignent la taille adulte à la fin de cette saison. Les limaces se reproduisent typiquement à la fin de l'été et meurent au cours de l'hiver suivant. Elles sont hermaphrodites.

La limace
Le corps est divisé en trois régions: la tête, la masse viscérale et le pied.
La tête bien développée, située à la partie antérieure du corps, porte deux longs tentacules rétractiles, au sommet desquels se trouvent les yeux. Les deux tentacules plus courts servent à l'olfaction, ils sont situés à proximité de la bouche ventrale.
La masse viscérale a la forme d'un manchon allongé. La partie antérieure et dorsale de cette masse est recouverte par un bouclier ovale : c'est le manteau. L'Arion est un Mollusque qui ne possède plus de coquille. Ou plus exactement, la coquille n'est plus représentée que par quelques granulations calcaires incluses dans le manteau. L'orifice que vous voyez à droite du corps est le pneumostome: il représente l'ouverture unique de la cavité palléale.
Le pied s'étend à la face ventrale de l'animal. Il est très musclé, et cilié et aide au déplacement.
5.2. Examen interne - 5.2.1. Le tégument
Passons à l'examen interne; le tégument se compose d'une seule assise épithéliale qui recouvre la totalité du corps. Il est très riche en cellules glandulaires, produisant le mucus; il est cilié dans certaines régions du corps, par exemple le pied.
5.2.2. Le système locomoteur
La musculature comporte de nombreux faisceaux musculaires qui doublent partout la face profonde du tégument. Un muscle longitudinal très épais forme le pied. Toute cette musculature permet à l'animal de ramper ou de se ramasser fortement sur lui-même lorsqu'il est menacé.
La limace se déplace par reptation: les muscles du pied se contractent par ondes successives d'arrière vers l'avant. Les cellules glandulaires du pied sécrètent une grande quantité de mucus qui permet à l'animal d'adhérer au support et de glisser sur un tapis liquide. On peut suivre une Limace à la trace, avec la traînée de mucus séché qu'elle laisse derrière elle. Cette trace est utilisée par certains insectes prédateurs pour localiser leur proie.
5.2.3. Le système digestif
L'Arion est essentiellement végétarien. Voici son système digestif. Nous observons de l’avant à l’arrière: la bouche avec la radula, le pharynx, l'oesophage court dans lequel débouchent deux glandes salivaires, le long estomac partiellement enveloppé d'une glande (ou caecum) digestive. Enfin, l'intestin s'enroule d'arrière en avant, en plusieurs spires autour de l'estomac pour déboucher auprès du pneumostome. C'est là, curieusement, que se situe également l'anus. Cette disposition particulière de l'intestin provient d'un double mouvement de flexion et de torsion qui s'est produit au stade larvaire, nous y reviendrons.
Faisons fonctionner ce système: la nourriture râpée par la radula arrive dans l'oesophage. Des enzymes sécrétés par les glandes salivaires y entament la digestion des glucides. Les glandes salivaires produisent également du mucus, qui lubrifie la radula et facilite le transit des particules ingérées. Ensuite, dans l'estomac, les aliments subissent un broyage mécanique sous l'action de la musculature et de la rotation d’une tige rigide de nature protéique, la tige cristalline. Ce broyage provoque une désintégration poussée des fragments végétaux. Toujours dans l'estomac, grâce aux enzymes libérés par l’abrasion du style cristallin, les aliments subissent une digestion extracellulaire. Les petites particules alimentaires assimilables sont emportées par des mouvements ciliaires vers le caecum digestif, où elles sont phagocytées. Donc, c'est dans cet organe que se produit la digestion intracellulaire et l'absorption. Les matériaux de trop grande taille et les déchets demeurent dans l'estomac où ils sont agglutinés et passent dans l'intestin, qui en absorbe l'eau et des sels minéraux avant de les évacuer par l'anus.
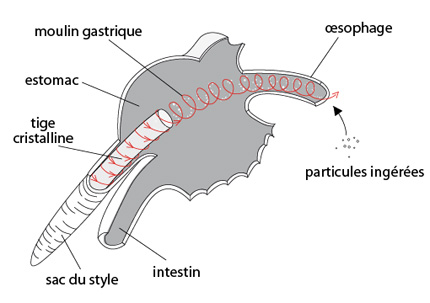
La tige cristalline
Cette tige cristalline présente chez les gastéropodes fait également partie du système digestif des mollusques monoplacophores et bivalves.

5.2.4. Le système respiratoire
Voyons maintenant le système respiratoire des pulmonés. Regardez d'abord en haut à gauche, le schéma de la dissection. Lorsqu'on découpe le manteau en suivant son pourtour, on voit que l'on peut le soulever comme un couvercle. Ce manteau délimite une poche close sur le dos de l'animal; c'est la cavité palléale, qui communique avec l'extérieur par le pneumostome. Le toit de la cavité palléale fonctionne comme un poumon. Près de cette cavité MAIS localisé dans la masse viscérale, nous trouvons le coeur ainsi que le rein.
Le toit de la cavité palléale est parcouru de réseaux de capillaires sanguins à paroi très fine, appelé plexus pulmonaire. Or, la cavité palléale est constamment remplie d'air qui entre par le pneumostome lorsque le plancher de la cavité palléale s’abaisse, et les échanges gazeux s'opèrent avec le réseau vasculaire. C'est donc l'équivalent d'un poumon doté d’un mécanisme ventilatoire. Le pneumostome peut s'ouvrir ou se fermer selon les besoins: quand l'atmosphère est sèche, il se contracte de manière à éviter une trop forte évaporation. En atmosphère humide par contre, il reste béant. Notez que le reste de la surface corporelle, toujours humide, participe aussi aux échanges gazeux.
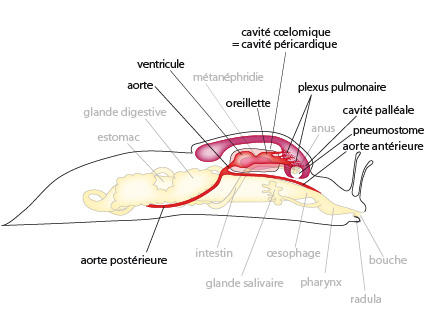
5.2.5. Le système circulatoire
Le coeur est situé antérieurement dans la masse viscérale. Il se compose d'une seule oreillette et d'un seul ventricule. Il est contenu dans un sac péricardique; ce sac représente un reste de la cavité coelomique.
L'oreillette reçoit l’hémolymphe des veines de la partie antérieure du toit de la cavité palléale où le sang s'est oxygéné. Puis le sang passe au ventricule, d'où il est propulsé dans l'aorte unique. Cette aorte se divise en 2 troncs antérieur et postérieur, irriguant respectivement la tête et le pied d'une part, et les viscères de l'animal d'autre part. Les aortes se ramifient puis se perdent dans des lacunes, sortes de cavités irrégulières que l'on observe dans tout le corps. Enfin, le sang est ramené au coeur par des ramifications veineuses après avoir traversé le plexus pulmonaire. C'est donc un système circulatoire ouvert: au cours de son trajet, le sang entre en contact direct avec les liquides interstitiels.

5.2.6. Le système nerveux
Le système nerveux de la limace est construit sur le modèle typique des protostomiens, avec des ganglions antérieurs reliés à une paire de cordons nerveux ventraux. En raison de la torsion qui se produit lors du développement, la double chaîne nerveuse forme un 8.
Voici des organes des sens chez la Limace. Les yeux situés au sommet des grands tentacules de l'Arion se composent des éléments suivants: la cornée externe, épiderme mince et transparent; la cornée interne située sous la précédente, et transparente elle aussi; une couche de cellules photoréceptrices et de cellules pigmentées qui forment la rétine. Les cellules photoréceptrices rejoignent leurs axones en un nerf optique qui se dirige vers le cerveau. Le cristallin sphérique et transparent occupe le centre de l'oeil. Ces yeux permettent à la limace de percevoir la lumière, le mouvement, et probablement de former une image rudimentaire.
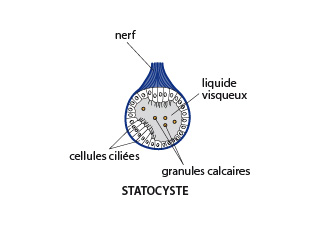
Le statocyste
Voici une coupe dans un statocyste. Ce sont des organes de l'équilibre, accolés au cerveau. La limace en possède deux; Ils se composent d'une vésicule creuse bordée de cellules ciliées, en rapport avec le nerf qui entoure l'organe. La vésicule est remplie d'un liquide visqueux dans lequel flottent de granules calcaires. Lors des déplacements de l'animal, les granules viennent heurter les cellules ciliées, qui transmettent au nerf les impulsions reçues. L'Arion est ainsi avertie de la position dans laquelle elle se trouve.
Les récepteurs olfactifs, gustatifs et du tact ne sont pas groupés en organes. Ce sont des cellules sensorielles dispersées, mais plus spécialement localisées dans la partie antérieure du corps.
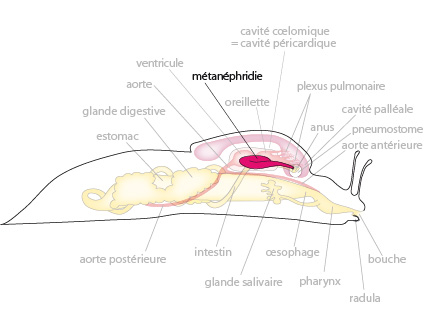
5.2.7. Système excréteur
Le système excréteur de la limace consiste en une seule métanéphridie, semblable à celles des annélides. A une extrémité, le nephrostome est ouvert dans la cavité péricardique où il récolte les déchets de l’hémolymphe revenant des différentes parties du corps; le néphridiopore est ouvert à l’extérieur au niveau de la cavité palléale (ici le pneumostome). Notez que les gastéropodes terrestres conservent l’eau en excrétant l’acide urique, peu soluble, et dont l’élimination emporte moins d’eau que dans le cas de l’ammoniaque. Cependant, ils sont capables de résister à de fortes déshydratations, causées notamment par la production de leur sillon muqueux.

5.2.8. Système reproducteur
Les Gastéropodes terrestres sont hermaphrodites. Un ovotestis, gonade produisant à la fois des gamètes mâles et femelles, est logé dans la partie postérieure de la Limace. De l'ovotestis part le canal hermaphrodite, long et mince, qui se poursuit par un canal plus large, l'ovospermiducte constitué de 2 gouttières. Ces deux gouttières se séparent ensuite en un spermiducte et un oviducte. Les limaces possèdent donc à la fois l'organe mâle et l'organe femelle, mais à cause d'une mécanique compliquée de leur copulation (sur laquelle nous ne nous étendrons pas), il y a heureusement fécondation croisée. Cependant, les accidents d'autofécondation ne sont pas rares.
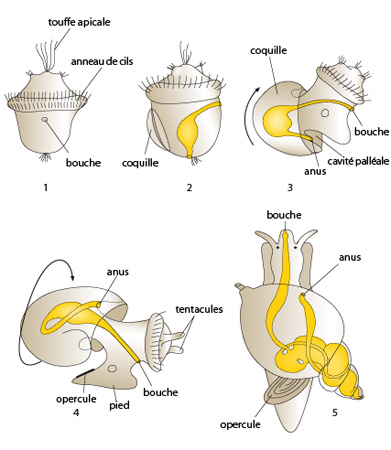
Les Gastéropodes
Chez les Gastéropodes primitifs, on retrouve la larve trochophore, très semblable à celle des Annélides. Cette larve possède une couronne ciliée équatoriale et une touffe apicale de cils. Quelques différences cependant: une glande coquillère se développe sur la surface dorsale et est divisée en sections transverses. Le pied se développe de l'autre côté et une paire d'yeux apparaît juste au-dessous de la couronne ciliée.
Chez les Gastéropodes marins, la larve la plus caractéristique est la larve Veligère: elle ressemble encore à une trochophore, mais présente un développement plus complexe. Voyons en détail ce développement. En (1) vous voyez une larve trochophore en vue ventrale avec une touffe apicale, un anneau de cils et sa bouche. Ensuite la larve trochophore (en 2, vue latérale droite) a acquéri une glande coquillère qui formera la coquille. Une flexion a lieu et nous retrouvons en (3) la larve véligère en vue latérale droite avec sa masse viscérale ayant subi un mouvement de flexion comme indiqué par la flèche et son tube digestif en forme de U. Finalement cette larve véligère subit une torsion de 180° comme indiqué sur le schéma en (4). Cette torsion des organes internes amène la cavité palléale avec les cténidies et l'anus à une position antéro-latérale au-dessus de la tête (5).
Comme vous le voyez, cette larve véligère possède une coquille, un pied et un opercule.
Une remarque importante à ajouter ici. Les gastéropodes prosobranches ont subi une torsion de 180° et ont leurs branchies en avant du corps comme indiqué dans ce schéma. Par contre les opisthobranches ont subi une torsion de seulement 90° et leurs branchies sont restées à l’arrière du corps. Certains gastéropodes terrestres ont acquis une poche respiratoire plutôt que des branchies. Ces gastéropodes, comme la limace, ont également subi une flexion et torsion de 180° lors de leur développement.

6. Exemples-type 3 : les Bivalves
Etudions maintenant la moule, Mytilus edulis, que vous verrez aussi lors des travaux pratiques au cours d’une dissection ou l’anodonte. Elles appartiennent à la classe des Bivalves ou Lamellibranches. Nous pourrons alors comparer les structures de la limace et de la moule, et leur adaptation à deux milieux très différents.
La moule mesure environ 5-6 cm. Sa coquille est formée de deux valves, calcaires et colorées extérieurement en brun par la conchioline, un complexe protéique servant de matrice pour le dépôt des cristaux de carbonate de calcium. Chaque valve montre des stries de croissance, qui correspondent à l'alternance des saisons favorables et défavorables.
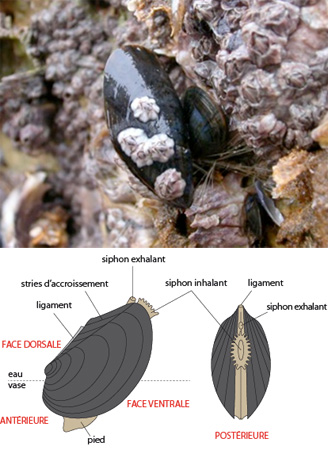
6.1. Examen externe
Ce schéma de gauche montre la position habituelle de la moule sur l’estran. Orientons l'animal. Ses deux valves symétriques s'articulent sur une charnière dorsale, constituée d'un ligament élastique. L'arc d'ouverture des deux parties de la coquille représente la face ventrale. Les siphons, dirigés vers le haut, indiquent la face postérieure, et à l'opposé, on trouve la face antérieure.
Si vous tournez la moule de manière à regarder sa partie postérieure, celle où se trouvent les siphons, vous voyez, comme l'indique le schéma de droite, que le siphon exhalant est plus dorsal que le siphon inhalant. Ces siphons servent à la circulation de l'eau, dont nous verrons toute l'importance lors de l'étude physiologique de l'animal.
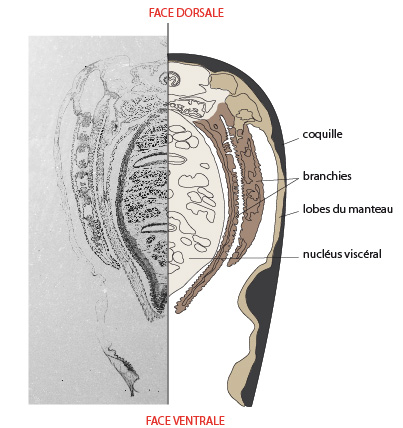
6.2. Examen interne
Voyons une coupe transversale. En noir, c'est la coquille formée de ses deux valves gauche et droite. En beige foncé, c'est le manteau qui recouvre le corps de l'Anodonte et qui secrète la coquille. En beige clair, c'est le nucleus viscéral se terminant par le pied à la face ventrale. Le nucleus viscéral contient les différents systèmes d'organes: digestif, circulatoire, excréteur, et génital. Les branchies, ici en marron, sont situées entre le nucleus viscéral et le manteau dans la cavité palléale.
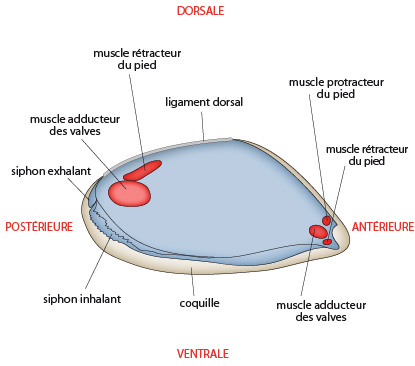
6.2. Examen interne
Si l'on détache la valve droite de la coquille, on voit en bleu, les lobes du manteau. Pour enlever la valve, il a fallu sectionner plusieurs muscles, représentés en rouge sur le manteau. Les deux gros muscles adducteurs des valves maintiennent les coquilles closes et servent d'antagonistes au ligament dorsal, qui tend à maintenir les valves ouvertes. Le ligament est composé de protéines non calcifiées. Les muscles adducteurs sont disposés transversalement d'une valve à l'autre. Les petits muscles rétracteur et protracteur permettent les mouvements du pied.

La moule
Voici l'intérieur de la coquie d'une moule.

La moule
Si l'on écarte les lobes du manteau, on aperçoit les branchies qui recouvrent le nucleus viscéral, dont on voit le pied en rouge. A la partie antérieure, la bouche est entourée de quatre palpes labiaux. On comprend maintenant pourquoi cette partie de l'animal est appelée antérieure. Cependant, contrairement à la Limace, la moule ne possède pas de tête distincte. A la partie postérieure, nous retrouvons les deux siphons.

Exercice :
Cette photo représente une moule ouverte comme vous le ferez lors de la dissection. Sur la photo, repérez les éléments que nous venons de décrire et orientez l'animal. Les schémas, c'est clair! La réalité est parfois moins évidente, mais c'est à elle que nous avons à faire, tout au long de notre vie professionnelle. Alors, observez, resituez les structures, et orientez l'animal.
6.2.1. Le tégument
Le système tégumentaire de l'Anodonte se compose du manteau et de la coquille. Le manteau est tapissé de deux épithéliums entre lesquels s'insère du tissu conjonctif de remplissage. L'épithélium dirigé vers la cavité palléale est constitué d'une seule couche de cellules ciliées; l'épithélium dirigé vers la coquille comporte des cellules glandulaires: certaines sécrètent la nacre, et d'autres assurent l'adhésion du manteau.
La coquille elle-même se compose de trois couches. De l'intérieur vers l'extérieur : la couche de nacre (ou hypoostracum) est finement lamellaire et cette constitution donne à la nacre son aspect irisé. L’hypoostracum/nacre est composé de calcite et de conchioline (une glycoprotéine); la couche prismatique ou ostracum est constituée de cristaux calcaires (CaCO3). Le périostracum est une fine couche de conchioline. La couche prismatique (ostracum) et le périostracum sont sécrétés par les bords du manteau uniquement. La coquille croît donc par son extrémité. Par contre, la nacre est déposée par toute la surface et recouvre donc l'intérieur de la coquille jusqu'à la ligne palléale. La présence de calcaire au niveau de l’hypoostracum et ostracum est facilement détectable en ajoutant de l'HCl à la surface. A l'opposé, aucune réaction ne sera observée en déposant du HCl sur le periostracum.

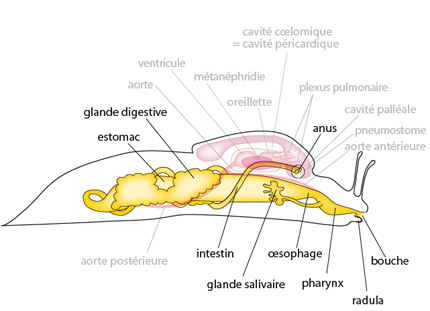
6.2.2. Le système digestif
La bouche, dépourvue de radula, conduit à un court oesophage qui mène les petites particules alimentaires dans l'estomac. L'estomac comporte un diverticule qui contient une tige cristalline animée d'un mouvement de rotation. Elle sert au malaxage des aliments. De plus, elle se dissout lentement par son extrémité libre, appendue dans l'estomac, en libérant des enzymes qui aident à la digestion extracellulaire.
La paroi de l'estomac est criblée d'orifices; ils donnent accès à l'ensemble des diverticules digestifs. Chez la moule, on observera l’hépatopancréas volumineux impliqués dans la synthèse d’enzymes digestives mais également dans la synthèse et dégradation des glucides ainsi que des processus de détoxification. Un long intestin fait suite à l'estomac. Il forme quelques circonvolutions avant de longer la face dorsale de l'animal, de traverser la cavité péricardique et le coeur (c'est étrange, mais c'est ainsi) et de se terminer à l'anus. L'anus s'ouvre dans la chambre cloacale du siphon exhalant (étant la partie dorsale de la cavité palléale). Les détritus alimentaires sont évacués à l’extérieur par le siphon exhalant.
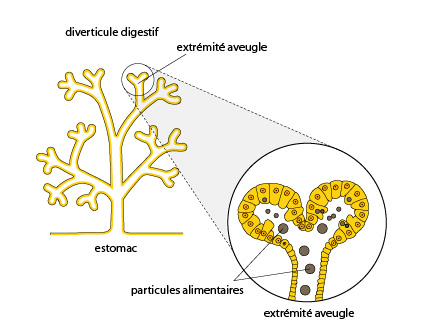
6.2.2. Le système digestif
Le schéma détaille les diverticules digestifs. Les particules alimentaires malaxées et partiellement décomposées dans l'estomac quittent celui-ci par ses orifices et sont acheminées dans les conduits ciliés, ramifiés, et en cul-de-sac. C'est dans ces diverticules que les particules alimentaires sont phagocytées. Là s'opère donc la digestion intracellulaire.
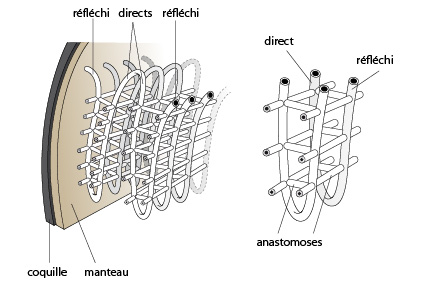
6.2.3. Le système respiratoire et circulatoire
Les organes respiratoires de l'Anodonte consistent en deux paires de branchies. Une de ces paires est détaillée ici. Chaque branchie se compose d'une série de filaments anastomosés entre eux, formant ainsi une grille. En coupe transversale, une paire de branchies dessine la lettre W.
Chaque filament est un tube creux dans lequel circule le sang. L'épithélium externe des filaments est cilié (les cils ne sont pas représentés). Le battement des cils engendre un courant d'eau à travers cette grille.
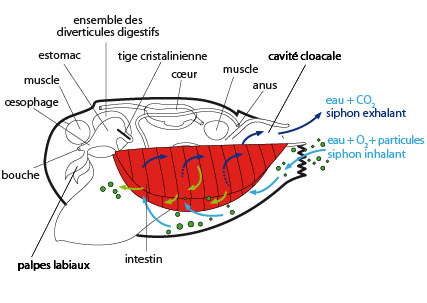
6.2.3. Le système respiratoire et circulatoire
Chargée d'oxygène et de particules diverses - grains de sable, protozoaires, bactéries, larves, fibres végétales, etc. - l'eau pénètre dans la cavité palléale par le siphon inhalant. Les flèches bleues indiquent le courant d'eau, et les flèches vertes indiquent le circuit des particules alimentaires.
Grâce au battement des cils des branchies, l'eau passe à travers la grille branchiale. Le sang qui circule dans les filaments capte l'oxygène et se débarrasse du CO2 dans l’eau circulant dans la cavité palléale puis expulsée par le siphon exhalant.
Les particules sont également entraînées dans la grille branchiale où elles sont triées. Les grosses particules denses tombent au fond de la cavité palléale. Lorsque les valves s'entrouvrent, elles sont rejetées dans l'eau. Les petites particules sont acheminées vers les palpes labiaux, grâce à l'action des cils.
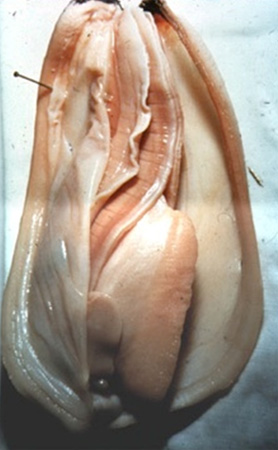
6.2.3. Le système respiratoire et circulatoire
Sur cette photo, récapitulons le rôle des branchies. Imaginons la moule dans la mer. Les branchies ciliées animent un courant d'eau qui entre par le siphon inhalant; on le voit en noir. L'eau chargée de particules et d'oxygène circule sur toute la longueur des branchies. Les aliments sont captés par les cils, triés et entraînés vers les palpes labiaux à la partie antérieure du corps.
En traversant les branchies, l'eau cède son oxygène, se charge en CO2 et ressort dans la mer par le siphon exhalant, que l'on ne voit pas ici, parce qu'il est caché par le lobe gauche du manteau.
Cette disposition des branchies en lamelles est caractéristique de toute une variété de Mollusques marins et d'eau douce, à coquille bivalve et sans tête distincte, que l'on range dans la classe des lamellibranches. La moule est un Mollusque Lamellibranche.
Les palpes labiaux sont plissés et ciliés. Ils effectuent un second tri des particules qui leur envoient les cils des branchies, selon leur grosseur et leur nature. Les éléments indésirables tombent dans la cavité palléale, et seront éliminés. Ceux qui constituent le régime alimentaire sont conduits vers la bouche. Les mécanismes de cette sélection étonnante sont encore inconnus; sans doute les palpes contiennent-ils des cellules spécialisées, chémoréceptrices.
Les branchies jouent également un rôle au niveau de l’osmorégulation.
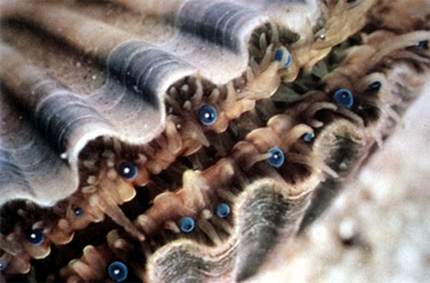
6.2.4. Le système nerveux
Chez les bivalves, comme chez ce Pecten (la coquille saint jacques), on note parfois la présence de nombreux petits yeux sur le bord du manteau, ou parfois sur les siphons chez d'autres espèces. Ces yeux peuvent détecter des changements de l'intensité lumineuse; ils comprennent aussi une cornée, une rétine et une lentille.
Le système nerveux est décentralisé. Le bord du manteau est tapissé de cellules nerveuses sensibles à la température, aux substances chimiques et à la lumière. Le système nerveux est réduit à trois paires de ganglions nerveux situés dans la partie antérieure, le pied et les viscères.
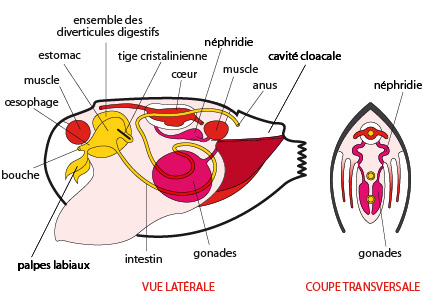
6.2.5. Le système reproducteur
La moule est dioïque: chaque animal possède une paire de gonades, soit des testicules chez les mâles et des ovaires chez les femelles. Chaque gonade est desservie par un conduit assez court qui aboutit, comme l'indique la coupe transversale, près de l'orifice excréteur des reins dans la cavité cloacale. Encombrés par leurs valves, ces animaux auraient quand même bien du mal à s'accoupler. Ce n'est pas nécessaire; voici comment la fécondation a lieu. Les femelles élaborent dans leurs ovaires des phéromones, qui passent dans l'eau ambiante et provoquent chez les mâles l'émission du sperme. Les spermatozoïdes sont évacués dans l'eau par le siphon exhalant, et se retrouvent dans la cavité palléale des femelles, en rentrant par leurs siphons inhalants. Ils y fécondent les ovocytes que la femelle a émis. Les zygotes se logent sur les lames des branchies externes, puis dans des poches de ces branchies, préparées pour les recevoir, et ils s'y développent dans le calme, avec une oxygénation idéale, pour qu'ils reçoivent constamment un courant d'eau fraîche.
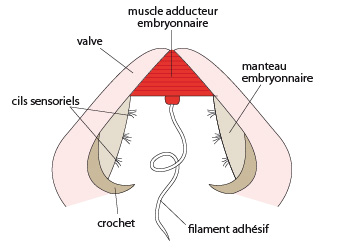
Glochidies
Chez certaines Bivalves d’eau douce, la larve ne ressemble plus du tout à une trochophore. Les oeufs produisent des larves appelées 'glochidies', mesurant un demi-millimètre. Elles comportent deux petites valves, chacune garnie d'un crochet, un manteau embryonnaire cilié, et un muscle adducteur portant un long filament adhésif. Pas de bouche, pas d'anus: le système digestif est quasi inexistant. La larve se nourrit par son manteau dont les cellules sont aptes à la phagocytose. Le crochet permet à la larve de s’accrocher aux branchies d’un poisson, de s’y développer avant de se détacher et descendre vers un substrat où elle s'établira.

7. Exemple-type 4 : les Céphalopodes - 7.1. Examen externe

Sepia officinalis
Voici un autre Mollusque marin : Sepia officinalis, communément appelé Seiche.
La Seiche vit non loin des côtes, sur les fonds sableux. C'est un prédateur très mobile. Elle peut si bien identifier son teint à celui du milieu qu'elle en devient très difficile à repérer. Son manteau peut varier sa coloration selon les teintes et les formes de son environnement (mimétisme) mais aussi en fonction de ses humeurs: belliqueuse, amoureuse ou effrayée. Elle est carnivore et se nourrit de Mollusques et de Crustacés.
Dans le corps de la Seiche, nous discernons 3 régions: la TETE, où vous voyez nettement un des deux yeux, sur le côté et un peu dorsalement, le PIED représenté par 8 bras ou tentacules courts et 2 plus longs, et la MASSE VISCERALE. La Seiche se range dans la Classe des Céphalopodes. Cette classe regroupe les Mollusques qui ont le pied sur la tête.
Observons cet animal plus attentivement. Sans tentacules, il mesure environ 40 centimètres. Les tentacules ont une forme pyramidale à trois faces. La face interne est complètement couverte de ventouses. Cette configuration permet à l'animal d'adhérer solidement au support, et d'agripper fermement ses proies.
Les deux longs tentacules, mesurant une vingtaine de centimètres, sont renflés à leur extrémité. Seul ce renflement est garni de ventouses. Ces tentacules peuvent se retirer à l'intérieur de poches appropriées. Ils ne sont pas utilisés pour la locomotion, mais seulement comme organes de préhension.
La masse viscérale est aplatie et entièrement recouverte d'un manteau, bordé d'une nageoire ondulante. En somme, le manteau a la forme d'un fourreau d'où sortent la tête et les tentacules.
La face dorsale offre une résistance dure car la Seiche possède une coquille calcaire mais elle est interne. De forme ovale et plate, elle est vendue par les oiseleurs sou sous le nom d'os de Seiche.
La coquille est sécrétée par le manteau. Primitivement externe, cette coquille est ensuite enfermée dans une invagination close du manteau. Elle est située sur toute la longueur de la face dorsale du corps. Il y a des coquilles de Seiche dans la salle. Vous pouvez en manipuler. La coquille est formée de minces lamelles calcaires soutenues par des piliers de la même matière. Cette architecture laisse des vides qui sont remplis de gaz. La coquille sert de flotteur.
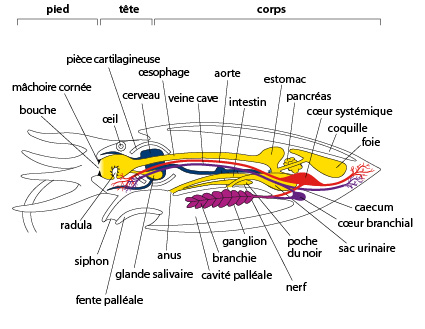
7.2. Examen interne
Chez la seiche nous distinguons 3 régions : le pied, qui contient les tentacules et le siphon, la tête et le corps (ou masse viscérale). Regardons d'abord la coupe sagittale du schéma. La coquille interne surmonte la masse viscérale, ventralement se situe la cavité palléale. Observez bien la situation de cette cavité chez la Seiche. Elle communique avec l'extérieur par la fente palléale. Un siphon s'engage dans l'ouverture de la cavité palléale. Il se situe sous la bouche, un peu plus bas que la couronne de tentacules.
La bouche, elle, s'ouvre au centre de la couronne de tentacules. Si l'on incise le manteau ventralement et que l'on rabat les volets formés par cette dissection, on constate que la cavité palléale est très vaste. Le siphon apparaît comme une hotte de cheminée, ou comme un entonnoir, dont l'ouverture la plus large débouche dans la cavité palléale. Les parois latérales et ventrales du siphon sont libres. Cependant, la paroi ventrale porte deux encoches: les boutonnières, dans lesquelles s'accrochent deux protubérances du manteau: les boutons. Cette curieuse attache, dite en bouton-pression maintient solidement le manteau au siphon et vice-versa. La seiche vivante n'ouvre jamais ses boutons.
Dans la cavité palléale, on aperçoit deux branchies et divers orifices: à savoir, l'anus, les 2 pores urinaires, et l'orifice génital qui n’est pas représenté sur ce schéma. Nous allons regarder cela plus en détail.
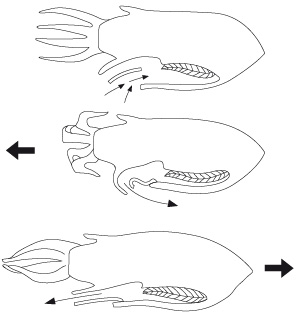
7.2.1. Système locomoteur
C'est assez compliqué une Seiche! Cette description externe va permettre de comprendre la locomotion.
Elle peut ramer en s'accrochant par ses tentacules au support, ou nager lentement en pleine eau grâce aux ondulations de sa nageoire. Mais elle peut aussi se servir du siphon pour se propulser rapidement, comme le montre la figure.
Quand l'animal relâche la partie ventrale de son manteau, l'eau s'engouffre par la fente palléale et remplit la cavité. Une contraction brusque du manteau augmente la pression de l'eau dans la cavité palléale, et force le pourtour intérieur du siphon à boucher la fente palléale. L'eau ne peut plus sortir que par le siphon. La force de l'eau quittant le siphon propulse l'animal vers l'arrière, comme une tuyère de réacteur.
Vous observerez également lors des travaux pratiques que le siphon est attaché à une série de muscles qui permettent d’orienter le siphon de gauche à droite.

7.2.2. Système digestif
Quand une Seiche voit une proie, elle la saisit et la porte à sa bouche. La cavité buccale contient deux mâchoires cornées et une radula. Deux glandes salivaires y déversent leurs sécrétions. Les glandes salivaires produisent un liquide permettant de paralyser les proies. Un long oesophage conduit la nourriture dans un estomac broyeur, très musclé, puis dans le caecum.
Le caecum reçoit les sucs digestifs de deux glandes appelées, un peu abusivement et par analogie foie et pancréas mais chez le calmar que vous verrez au TP on retrouve une glande digestive, l’hépatopancréas. La glande digestive a trois fonctions. La première est de sécréter des enzymes digestifs. La deuxième est d'accumuler des substances de réserve amenées par le système circulatoire, tels des sucres et des graisses. Ces réserves sont libérées massivement lorsque la Seiche a besoin d'une grande quantité de substances énergétiques en un temps très court. Ceci arrive, par exemple, lorsque l'animal fuit un prédateur, ou au contraire lorsqu'il poursuit une proie. La troisième fonction de cette glande est une fonction d'excrétion. Elle transforme les substances de déchets en ammoniaque, urée et acide urique. Ces produits passent alors dans le sang, d'où ils seront extraits par les sacs urinaires.
La digestion extracellulaire commence déjà dans la cavité buccale. Elle se poursuit dans l'estomac, mécaniquement, et s'achève chimiquement dans le caecum.
Les éléments indésirables sont conduits dans l'intestin qui débouche à l'anus, dans la partie antérieure de la cavité palléale. Puis les déchets sont éjectés dans la mer. C’est au niveau de l’intestin que se fait la réabsorption.
La poche du noir se jette dans l'intestin tout près de l'anus. C'est un organe propre aux Céphalopodes. La poche du noir produit un pigment: des grains de mélanine tenus en suspension dans un liquide. C'est l'encre que l'animal éjecte pour former autour de lui un nuage noir protecteur qui masque sa fuite, quand il veut échapper à un prédateur. L'encre sort par l'anus, puis le siphon. Jadis, on utilisait ce produit comme encre de plume, et actuellement, on l'utilise encore comme encre à dessin. Selon sa dilution, elle donne des teintes noires, brunes, vertes et finalement roses.
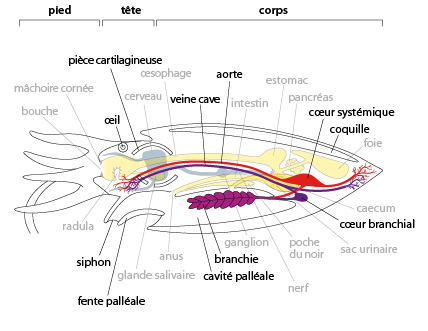
7.2.3. Système respiratoire
Le système respiratoire de la seiche comprend des branchies, organes flexibles et composés de filaments creux dans lesquels circule le sang. Elles baignent dans l'eau toujours renouvelée dans la cavité palléale. Le sang y fixe l'oxygène grâce à l'hémocyanine et s'y débarrasse de son CO2.
La limace possédait un système circulatoire ouvert. Mais par contre, chez la Seiche, le système circulatoire est fermé! Tout au long de son trajet, le sang est conduit dans des vaisseaux bordés d'un endothélium, et ne se déverse plus dans des espaces lacunaires. Il s'y ajoute, en outre, deux petits cœurs annexes qui pompent le sang dans les branchies.
Ce système fermé assure une pression sanguine plus importante et par conséquent, une circulation plus rapide. C'est nécessaire chez un animal actif qui doit pouvoir disposer par moments d'une grande quantité d'énergie.
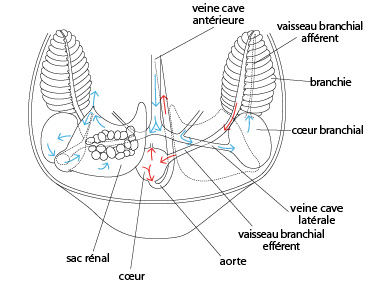
Le système circulatoire
Le cœur systémique se contracte et envoie le sang oxygéné aux différents organes du céphalopode via deux aortes (antérieure et postérieure). Le sang désoxygéné revient au niveau de deux cœurs branchiaux chacun relié à une branchie. En se contractant, un cœur branchial envoie le sang désoxygéné au niveau de la branchie via le vaisseau branchial afférent. Le CO2 peut ainsi être évacué. En repassant par le vaisseau branchiale efférent, le sang se recharge en oxygène. Le sang oxygéné arrive enfin au cœur systémique et la boucle recommence.
Après avoir irrigué les différents organes du corps, le sang revient par la veine cave. Cette veine traverse le sac urinaire. Là, le sang se débarrasse de ses déchets. C'est une particularité du système excréteur des Céphalopodes.
Vous remarquez que le mode de vie des différents Mollusques influence profondément la disposition d'organes que l'on pourrait qualifier de domestiques. C'est tout aussi net pour le système nerveux et les organes des sens.
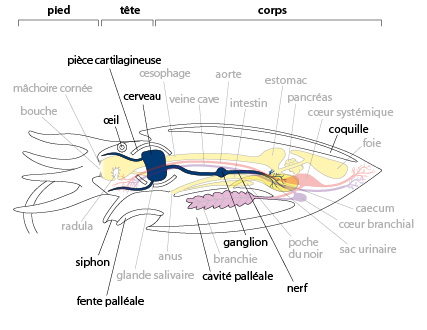
7.2.4. Système nerveux
Le système nerveux et les organes des sens de la Seiche et de tous les autres Céphalopodes sont bien plus performants que ceux des autres Mollusques.
Le volume du tissu nerveux est plus important et sa structuration est plus complexe. Des ganglions nerveux sont concentrés autour de l'œsophage, en une grosse masse qui forme un volumineux cerveau, d'où partent des nerfs se rendant à tous les organes du corps. Sur le trajet des nerfs, de nombreux ganglions s'individualisent; ils sont sans correspondant chez les autres Mollusques. Le cerveau est protégé dans une capsule de pseudo-cartilage qui forme une espèce de crâne.
Pour l'équilibre, la Seiche dispose de deux statocystes logés dans les pseudo-cartilages céphaliques, près du cerveau. Des cellules chémoréceptrices localisées dans la bouche la renseignent sur la nature de ses aliments. De nombreuses cellules sensorielles dispersées dans tout l'épiderme rendent l'animal très sensible au toucher.
Mais le plus spectaculaire, c'est la vue de la Seiche, son sens le plus développé.

L'oeil de seiche
Regardez cet œil de Seiche. Sa structure n'est-elle pas très proche de celle de nos yeux ?
En coupe, l'œil est plus ou moins sphérique. La partie antérieure est limitée par la cornée transparente. Elle est prolongée par une paupière circulaire formée par un repli de l'épiderme. Le diamètre de la paupière peut se réduire par la contraction d'un muscle logé dans ce repli. Derrière la cornée, on trouve l'iris. L'iris est un diaphragme: par augmentation ou diminution de son diamètre, il règle la quantité de lumière qui pénètre dans l'oeil.
Derrière l'iris, on trouve le cristallin: c'est une grosse lentille biconvexe maintenue par des supports musculaires. Le cristallin divise l'oeil en deux chambres: la chambre antérieure et la chambre postérieure, cette dernière est tapissée par la rétine. Chaque cellule rétinienne envoie une fibre nerveuse au ganglion optique, un relais sur la voie de communication avec le cerveau, par le nerf optique.
Le ganglion optique est protégé par des pièces de cartilage. On sait que l'œil de Seiche forme une image, et que cette image est droite. On sait aussi que cet œil détecte les couleurs.
On le voit, cette structure ressemble à celle de l'œil des Vertébrés. Les Mollusques et les Vertébrés sont deux groupes éloignés l'un de l'autre; ils n'ont pas d'ancêtres communs possédant cette structure oculaire. Ces yeux semblables, exerçant une fonction analogue, ont dû apparaître indépendamment au terme de ces lignées évolutives distinctes; on dit qu'il y a évolution convergente.
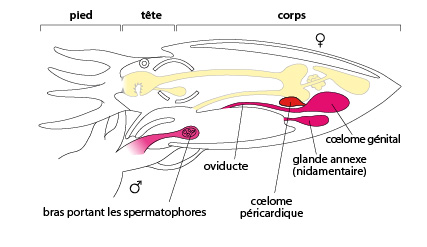
7.2.5. Système reproducteur
Les Céphalopodes sont dioïques. Chez le mâle, un des huit (ou dix) tentacules se modifie au moment de la reproduction: il se dilate, les ventouses rétrécissent. Ce tentacule, appelé hectocotyle, forme une sorte de cuillère qui sert à recueillir ses propres spermatophores, et à les introduire dans le corps de sa femelle. En effet, les spermatozoïdes sont empaquetés dans une enveloppe qu'on appelle spermatophore. Lors de l'accouplement, le mâle dépose ses spermatophores dans la cavité palléale de la femelle, où ils éclatent. Chez la femelle, la gonade se situe dans le coelome génital. Les oeufs descendent l'oviducte. Arrivés dans la cavité palléale, ils sont fécondés, et entourés individuellement d'une coque protectrice sécrétée par deux grosses glandes, appelées nidamentaires. Puis, ces œufs sont colorés en noir par l'encre de la poche du noir.

Les oeufs de seiche
La femelle recueille les œufs, et les attache soigneusement aux objets immergés en grappes compactes. C'est ce que l'on appelle les raisins en mer.
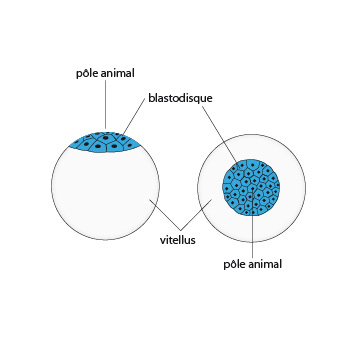
Les oeufs de seiche
Chaque œuf contient beaucoup de vitellus. Son diamètre est de 1 centimètre et demi environ. Par conséquent, la segmentation de l’oeuf est particulière :
Seul le pôle animal se fragmente en blastomères. Le pôle végétatif bourré de vitellus ne se divise pas. Comme résultat, on trouve au pôle animal un disque embryonnaire composé d'une seule couche de cellules, qu'on appelle blastodisque. C'est la première fois que nous voyons ce type de segmentation; il nous intéresse: nous le retrouverons chez les Reptiles et les Oiseaux. Passons la suite ... le développement continue dans la coque. En sortant de l'œuf, la jeune Seiche aura la même forme que l'adulte, elle est immédiatement capable de nager et de se nourrir.
De nombreux autres Mollusques montrent une segmentation plus typique, qui n'est pas troublée par la masse vitelline. Et alors, cette segmentation est très révélatrice: c'est une segmentation spirale, comme celle des Annélides !
Exercice :
Maintenant un exercice de synthèse: à partir des 5 espèces étudiées, pouvez-vous dégager les caractères essentiels qui définissent les Mollusques.

8. Exemple-type 5 : les Scaphopodes
Une toute petite classe: celle des SCAPHOPODES. Le survol succinct que nous leur consacrons n'est pas gratuit. Il a sa raison d'être, vous le verrez bientôt.
Autrefois, les espèces de ce groupe étaient appelées dents d'éléphant ou dents marines. Une coquille de Dentale est ouverte à ses deux extrémités, une large et une étroite. Quand la coquille est occupée par le Mollusque qui l'a sécrétée, l'extrémité large est habituellement enfoncée dans le sable, et l'ensemble s'incline vers le haut, laissant dépasser l'extrémité supérieure, étroite. Par la petite ouverture, l'eau entre dans la coquille et en sort; ce courant d'eau assure la fonction respiratoire. Le Dentale n'a pas de branchies; il respire par la surface de son manteau.

Le Dentale
Par l'ouverture la plus large de sa coquille, le Dentale étend son pied qui ressemble à un sabot de cheval et qu'il utilise pour fouir le sédiment. Derrière le pied, sur le bord du manteau presque cylindrique, un bouquet de tentacules ciliés entoure la bouche. Ils lui apportent les particules alimentaires: les petites sont avalées directement, les plus grosses sont râpées par la radula. Le tube digestif est replié en U et s'ouvre par un anus vers l'avant.
9. Origine, diversité et évolution - Origine du groupe
Les premiers fossiles sont antérieurs au début de l'ère Primaire (ou Paléozoïque). Les espèces fossiles sont nombreuses et illustrent l'exceptionnelle radiation adaptative du groupe. Leur abondance et la présence d’une coquille minéralisée facilitant la fossilisation, permettent leur utilisation comme guide stratigraphique par les géologues. Parmi, les plus célèbres, les Ammonites ont hanté les océans du Paléozoïque et du Mésozoïque, certaines dépassant les 2m de diamètre. Elles n'ont pas survécu à l'extinction du Crétacé. Malgré l'abondance de ces documents paléontologiques, les relations phylogénétiques entre les taxons fait toujours l’objet de débats intenses au sein de la communauté des biologistes. Depuis le Primaire, les Mollusques se sont diversifiés en de nombreux rameaux, dont beaucoup se sont éteints. Actuellement, on les trouve dans tous les milieux : marins, dulçaquicoles et terrestres.
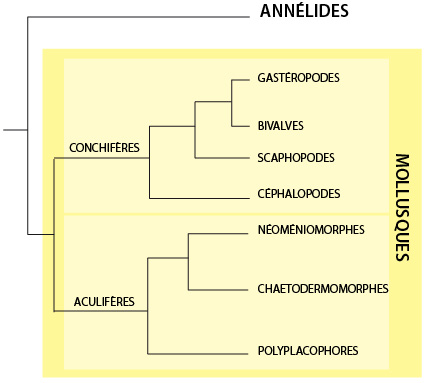
Relations phylogénétiques
Une étude récente dans Nature (Kocot et al., 2011) basée sur l’analyse de plus de 300 gènes a proposé cet arbre phylogénétique. Les groupes identifiés par leurs caractéristiques morphologiques y sont clairement identifiés. Les Mollusques se seraient séparés d’un ancêtre commun avec les annélides. Deux lignées auraient émergé: d’une part les aculifères, qui rassemblent les aplacophores (deux groupes de mollusques vermiformes) et les polyplacophores, et les conchifères, regroupant les gastéropodes, les bivalves, les scaphopodes et les céphalopodes. Ces derniers comprennent les poulpes, seiches et ammonites. Les coquilles des conchifères et les écailles, appelées sclérites, des aculifères ne seraient pas homologues.
La filiation des mollusques avec les annélides peut être décelée dans le développement de nombreux Mollusques actuels, dont l’œuf se segmente de la même manière que celui des Annélides, par segmentation spirale. De plus, les larves libres de certains Mollusques sont en fait des trochophores modifiées. Or, quand on retrouve dans les premiers stades de l'ontogenèse des deux groupes, des processus très semblables, on peut supposer qu'ils ont une origine commune.
Si les Annélides ont inventé la métamérisation, les Mollusques en ont apparemment perdu toute trace. Pas tous cependant. Découvert en 1942, lors d'une expédition océanographique dans le Pacifique, Neopilina a suscité un très grand intérêt. On connaissait des coquilles fossiles de la même Classe, celles des Monoplacophores, datant du Cambrien et du Dénovien. Aucun spécimen vivant de ce groupe n'était connu auparavant... Chez Neopilina, comme chez les autres Mollusques, aucune métamérisation grossière n'est néanmoins apparente: il possède une coquille simple et un pied non segmenté.

Neopilina
Par contre, à l'examen interne, Neopilina montre une répétition de divers organes pouvant faire penser à un vestige de métamérisation: deux paires d'oreillettes, deux paires de gonades, 8 paires de muscles rétracteurs du pied, 5 paires de branchies, 6 paires de néphridies. Cette disposition apparemment métamérisée a amené certains à l’assimiler à la conservation d’un caractère primitif d’un ancêtre qui aurait pu être un annélide. Cependant, les analyses génétiques n’ont pas pu, à ce jour, placer avec certitude cet animal unique, qui pourrait avoir acquis sa métamérisation secondairement. Néanmoins l’étude en 2006 de Giribet et al. dans PNAS a analysé un specimen de monoplacophore et démontre que ce groupe forme un clade avec les polyplacophores.
Diversité
Les mollusques constituent un des embranchements les plus diversifiés, avec plus de 100.000 espèces actuelles décrites et 35.000 espèces fossiles disparues.
Au sein du groupe aplacophor on retrouve 2 clades de mollusques vermiformes peu connues: les Chaetodermomorphes ou caudofovéates et les solénogastres ou Néoméniomorphes.
Les caudofovéates
Les caudofovéates sont des mollusques fouisseurs qui contiennent dans la partie postérieure du corps des cténidies, ils ont également une radula. Ils vivent dans les sédiments du milieu marin et il y a environ 100 espèces décrites à ce jour. Le manteau recouvre entièrement le corps secrétant des écailles calcaires.

Les Solénogastres ou Néoméniomorphes
Les Solénogastres ou Néoméniomorphes sont une classe de mollusques en forme de vers. On les rencontre dans toutes les mers du monde. Ils sont benthiques et vivent généralement sur les plantes ou les animaux fixés sur les fonds marins. Ils ont également une radula et des branchies. A ce jour il y a environ 350 espèces décrites.

Les polyplacophores
Les polyplacophores sont des mollusques brouteurs, environ 900 espèces décrites. Ils sont benthiques et souvent retrouvés dans les zones littorales. Ils se nourrissent d'algues microscopiques poussant sur les substrats durs qu’ils arrivent à brouter grâce à leur radula. Ils ont une forme aplatie ; leurs plaques dorsales sont constituées d'aragonite multicolore, décorée de motifs, lisse ou sculptée. Leur reproduction sexuée est typique des mollusques avec une fécondation externe et une embryogenèse qui se déroule dans l'eau de mer pour donner une larve planctonique trochophore. La larve se dirige ensuite vers les fonds marins, perd ses cils et devient un juvénile benthique.