Les spongiaires
Les Spongiaires sont le résultat d'un des passages de l'état unicellulaire à l'état pluricellulaire. L'état pluricellulaire permet une augmentation de taille des individus et le développement de structures complexes variées. Apparemment toutefois, cet essai n'a pas eu un succès remarquable; les Spongiaires n'ont pas donné d'autre ligne de descendance. Pour souligner ceci, certains zoologistes les rangent dans le règne des Parazoaires, par opposition aux autres Métazoaires.
Fiche récapitulative
- Aussi appelés Porifères
- Métazoaires dits « Parazoaires », c-à-d « à côté » des Métazoaires vrais, parce qu’ils ne comportent pas de vrais feuillets embryonnaires (ectoderme, mésoderme, endoderme), dont nous commencerons à parler avec les Cnidaires.
- Marins ou dulçaquicoles
- Solitaires ou coloniaux
- Environ 5500 espèces décrites
- Fixés à un support
- Absence d’organes vrais,
- Occurrence de cellules flagellées, les choanocytes, faisant circuler le milieu aquatique à travers de canaux ouverts sur l’extérieur
- Fonctionnement des cellules peu ou pas coordonné,
- Reproduction sexuée (le plus souvent hermaphrodisme et fertilisation croisée) et asexuée (bourgeonnement).

1. Présentation du groupe
Nous allons rapidement passer en revue l'Embranchement des Spongiaires. Il regroupe les plus primitifs des animaux pluricellulaires. En voici un exemple : cette éponge de mer colonise un rocher.
Le passage de l'état unicellulaire à l'état pluricellulaire est une étape cruciale de l'évolution. L'animal échappe ainsi aux limitations de la cellule isolée, il peut augmenter de taille et édifier des variations architecturales plus complexes, irréalisables dans l'état unicellulaire.
Tous les Spongiaires sont des animaux aquatiques. Excepté quelques espèces d'eau douce, les éponges vivent en milieu marin. Comme dans l'exemple ici, les spongiaires se fixent sur différents supports solides comme des rochers, des coquillages, etc. ; cependant, il en existe aussi sur des substrats moins fermes, comme du sable ou sur des fonds vaseux. Ils demeurent fixés pendant leur vie adulte ; on qualifie ce mode de vie de sessile.

La forme d'une éponge
Certaines éponges ont approximativement la taille d'un grain de riz, d'autres par contre mesurent plus d'un mètre. Quelques-unes ont une symétrie rayonnante, mais la majorité d’entre elles présentent une forme très irrégulière. Elles peuvent être massives, dressées sur leur support, arborescentes ou encroûtantes (c-à-d formant une croûte à la surface de leur substrat). La forme qu'adopte une espèce donnée est fortement influencée par les caractéristiques abiotiques de leur environnement telles que l'inclinaison du support, l'espace disponible, la vitesse et la direction du courant d'eau qui la baigne. Ainsi, une même espèce peut se présenter sous différentes apparences et la forme d’une éponge n’est pas un caractère morphologique informatif pour déterminer son statut taxonomique.
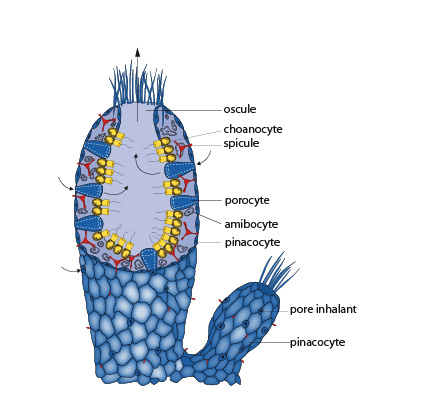
2. Exemples-type - 2.1. Examen externe
Pour comprendre la structure de base et la diversité cellulaire des éponges, nous commencerons par examiner une forme primitive.
La forme fondamentale s'observe le mieux chez l'individu jeune et solitaire, qui présente une structure de type « ascon » en forme de tube. Une double paroi compose ce tube : l'ectoderme extérieur ou pinacoderme formé par les pinacocytes, et l'endoderme interne ou choanoderme formé par les choanocytes. La surface de l'éponge est perforée de nombreux petits orifices, les pores inhalants. Ils donnent accès à une cavité centrale (l’atrium) qui débouche à l'extérieur par une large ouverture, l'oscule, située au sommet du tube.
Un courant d'eau traverse ainsi constamment l'éponge, l'eau entrant par les pores inhalants, baigne la cavité centrale puis sort par l'oscule.

2.2. Examen interne - 2.2.1. La paroi - Types cellulaires
Une nomenclature existe définissant les différents types cellulaires de l'éponge. Cette nomenclature est complexe. Nous vous en présentons un aperçu afin que vous puissiez vous rendre compte de la diversification des formes et des fonctions des cellules d’éponge.
Des cellules aplaties quelque peu contractiles, les pinacocytes, forment la surface extérieure du corps. Cette couche de cellules n'est pas un véritable épithélium, parce qu'elle ne repose pas sur une lame basale comme chez les autres métazoaires. Les pinacocytes à la base de l'éponge sécrètent un matériau qui fixe l'animal au support.
Chaque pore inhalant est entouré d'une seule cellule, le porocyte. Par contraction, celui-ci peut ouvrir ou fermer le pore.
Le diamètre de l'oscule peut aussi varier : il contrôle la vitesse du flux d'eau et parfois l'arrête totalement. Chez certaines éponges, ce contrôle est facilité par un type particulier de cellules appelées myocytes. Ces myocytes montrent quelques similitudes avec des cellules musculaires lisses des métazoaires vrais (« Eumétazoaires »). Elles ont une forme semblable et sont contractiles.
Sous les pinacocytes se trouve une matrice de protéines gélatineuses, la mésoglée, qui contient des cellules amiboïdes et le matériel squelettique. Les pinacocytes, la mésoglée et les cellules qui la peuplent forment la couche extérieure et moyenne du tube parfois nommé l'ectomésenchyme.
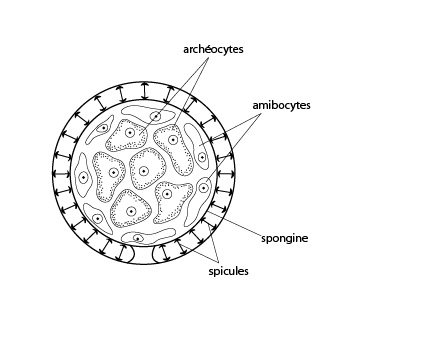
Les archéocytes
Parmi les cellules amiboïdes, celles qui comportent un gros noyau sont les archéocytes. Elles sont phagocytaires et jouent un rôle dans la digestion ; quand les archéocytes se limitent à ce rôle, on les dénomme amibocytes, mais les archéocytes sont capables de former d'autres types cellulaires quand le besoin s'en fait sentir. De telles cellules qui peuvent se différencier en d'autres types cellulaires sont qualifiées de toti/multi-potentes. En général, chez les Eumétazoaires et les Vertébrés en particulier, on ne trouve plus de cellules totipotentes au-delà des premières cellules embryonnaires.
Les cellules aux longs prolongements cytoplasmiques sont appelées collencytes; elles sécrètent des fibres collagènes dispersées. Ces fibres ont un rôle de soutien dans l'animal. Beaucoup d'éponges possèdent des lophocytes mobiles qui sécrètent aussi des fibres collagènes.

La spongine
En puis enfin, les spongiocytes fabriquent des protéines fibreuses proches du collagène, la spongine. Le squelette de spongine est organisé comme un réseau de fibres interconnectées d'épaisseur variable. Cette photo prise au microscope nous montre des fibres de spongine isolées, colorées en rouge.
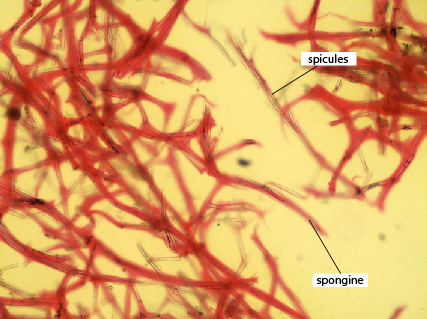
Fibres de spongine
Cette photo-ci montre également des fibres de spongine, accompagnées de spicules, qui sont ces sortes d’aiguilles claires plaquées sur la spongine.
La forme de l’éponge est renforcée par un squelette présent dans la mésoglée. Il peut être constitué de spicules, structures très effilées et fortement minéralisées. Dans notre exemple, elles sont constituées de carbonate de calcium. Ces épines constituent aussi un système anti-prédation, infligeant des blessures aux prédateurs qui tenteraient d’ingérer des tissus de l’éponge.
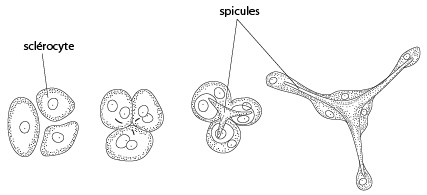
Les spicules
Le squelette de spicules est sécrété par les cellules sclérocytes. En général, pour fabriquer un seul spicule, plusieurs sclérocytes interviennent et le processus est relativement compliqué. Prenons par exemple la formation d'un spicule calcaire à trois branches. Trois sclérocytes s'associent, et chaque membre du trio se divise alors en deux. Une branche de spicule est sécrétée entre chaque paire de cellules filles. Les trois branches fusionnent à leur base. Chacune des trois paires de sclérocytes se déplace le long de chaque rayon. Une cellule sécrète la pointe et l'autre épaissit la base du spicule, qui correspond à un cristal de calcite ou d’aragonite.
Les spicules peuvent avoir des formes très diverses, mais leur morphologie est assez caractéristique d’une espèce et servent pour cette raison de critère taxonomique.
Les choanocytes
Enfin, la cavité centrale est bordée de cellules choanocytes. Ces choanocytes ressemblent de manière frappante aux Protistes Choanoflagellés. Nous en reparlerons. Le choanocyte est ovoïde, et l'extrémité tournée vers la cavité centrale porte un flagelle entouré d'une collerette percée de fentes. Les choanocytes sont responsables du mouvement de l'eau à travers l'éponge, grâce au battement de leur flagelle. Ils participent aussi à la digestion et à la reproduction sexuée.
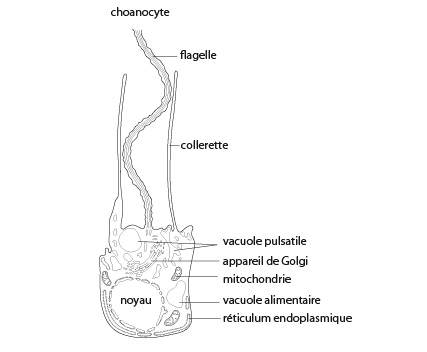
Structure d'un choanocyte
La figure est une représentation d'un choanocyte tel que vu en microscopie électronique.
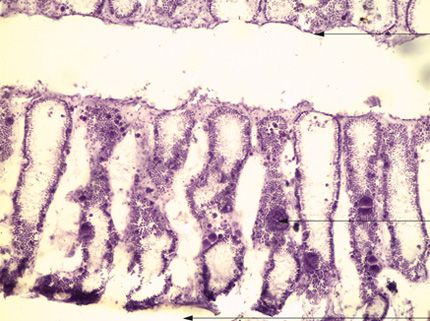
Grantia
Voici la photographie d'une coupe de l’éponge Grantia. Tentez d'y repérer les différents éléments, pour fixer l'organisation qui vient d'être décrite.
2.2.2. La locomotion
Les éponges adultes sont incapables de quelque locomotion que ce soit, cependant, certaines espèces peuvent se contracter ou modifier un peu la forme de leur corps.
2.2.3. Système digestif
La physiologie de l'éponge est largement dépendante du courant d'eau qui la traverse. L'eau apporte l'oxygène et la nourriture, puis emporte les déchets de la respiration et de la digestion.
Le volume de l'eau pompée par une éponge est remarquable. On a pu l'évaluer à 22.5 litres/jour pour un individu de 10 cm de haut et de 1 cm de diamètre. Le courant est produit par le battement des flagelles des choanocytes, mais il n'y a pas de coordination ni de synchronisation dans ces battements. C'est la configuration des tapis de choanocytes qui engendre et oriente le courant.
Les éponges se nourrissent de particules organiques extrêmement ténues, ce qui les classe parmi les animaux dits microphages. En effet, seules de très petites particules peuvent rentrer par les pores inhalants dont la taille peut descendre jusque 5 µm. Elles sont finalement capturées par les choanocytes lors du passage de l'eau au travers des microvillosités de la collerette, puis phagocytées par le choanocyte. L’espace entre deux microvillosités de la collerette est de l’ordre de 0,1-0,2 µm, ce qui permet de retenir des bactéries, des protistes,... Le choanocyte peut transférer la particule engloutie à d'autres cellules, les amibocytes par exemple. Ces derniers paraissent être les principaux sites de digestion. Que ce soit dans le choanocyte ou dans l'amibocyte, la digestion est intracellulaire, à l'intérieur de vacuoles alimentaires (voyez le schéma du choanocyte vu auparavant). Les amibocytes sont aussi des centres de stockage des réserves nutritives.
2.2.4. Système respiratoire - 2.2.5. Système circulatoire - 2.2.6. Système excréteur
L'architecture de l'éponge et le flux d'eau constant qui la traverse impliquent que la plupart des cellules sont en contact direct avec le milieu extérieur. L'échange des gaz O2 et CO2 se produit par simple diffusion entre le courant d'eau et les cellules. Quant aux résidus inassimilables de la digestion éjectés hors des cellules et les déchets azotés du métabolisme, ils quittent le corps avec le courant d'eau exhalé. La plupart des cellules des éponges d'eau douce possèdent une ou plusieurs vacuoles pulsatiles qui constituent le principal système osmorégulateur de ces animaux, évacuant continuellement l’eau qui s’infiltre dans les tissus.
2.2.7. Système nerveux
On n'a pas trouvé de système nerveux chez les éponges et les réactions aux stimulations extérieures restent très localisées. La faible coordination entre cellules est assurée par la transmission des molécules chimiques messagères diffusant dans la paroi et par le déplacement des cellules amiboïdes.
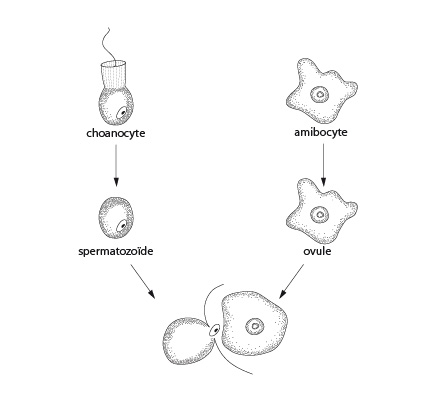
2.2.8. Système reproducteur - La reproduction sexuée
Les éponges sont en général hermaphrodites mais elles produisent leurs gamètes à des moments différents. C’est ce qu’on appelle une fécondation croisée. La reproduction sexuée et l'embryologie des éponges sont très particulières. Très bizarrement, des spermatozoïdes sont produits à partir de choanocytes. Certains choanocytes perdent leur collerette et leur flagelle et se transforment en spermatogonies. Nous assistons donc ici à un cas de dé-différenciation suivi d'une différenciation dans une autre voie. Chez d'autres espèces, les spermatogonies se différencient directement à partir d'archéocytes. Les spermatogonies se multiplient, subissent une méiose, maturent en spermatozoïdes et quittent l'éponge-père grâce au courant exhalant. Par ailleurs, des archéocytes ou amibocytes accumulent des réserves nutritives en dévorant quelques cellules adjacentes et se différencient en ovules. Les ovules restent situés dans la mésoglée et attendent qu'un spermatozoïde venant d'une autre éponge soit amené par le courant inhalant.
La fécondation est aussi très particulière : lorsque ce spermatozoïde a atteint une chambre à choanocytes, il pénètre un de ces choanocytes. Les deux cellules perdent leur flagelle et entrent dans la mésoglée. Elles s'y promènent jusqu'à rencontrer un ovule. Le choanocyte transporteur fusionne avec l'ovule et y transfère le spermatozoïde. La fécondation peut alors avoir lieu. Dans la majorité des espèces, le développement jusqu'au stade larvaire se produit à l'intérieur du parent.
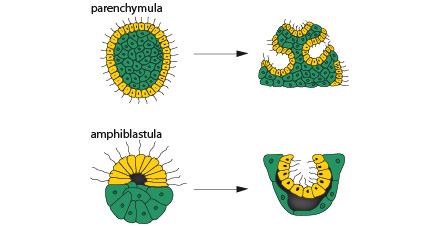
Le zygote
Chez certaines éponges, le zygote se divise de manière complète et mène à un stade larvaire appelé parenchymula. Des cellules flagellées couvrent toute sa surface externe. L'intérieur de cette larve contient la plupart des types cellulaires de l'adulte sauf des choanocytes. Cette petite larve brise la paroi du corps de son parent, et mène une brève existence libre.
D’autres éponges se développent sous la forme d’une autre larve, dénommée amphiblastula. Celle-ci est creuse. Un de ses hémisphères est composé de petites cellules flagellées, l'autre de grosses cellules non flagellées.
Après s'être fixées, ces deux larves subissent une réorganisation interne importante comparable à la « gastrulation » typique du développement d'autres animaux.
Chez la parenchymula, les cellules externes flagellées perdent leur flagelle et se déplacent vers l'intérieur de la larve où elles forment les choanocytes. Les cellules internes quant à elles se déplacent à la périphérie et se différencient en pinacocytes. Donc, les choanocytes sont formés par les cellules externes et les pinacocytes par les cellules internes. Vous comprenez la difficulté de parler ici de cellules dérivées de l'ectoderme et de l'endoderme, comme nous le ferons pour les Eumétazoaires.
Chez l'amphiblastula, la réorganisation qui suit la fixation se produit généralement par invagination: les petites cellules flagellées deviennent internes et les grosses cellules finissent par les entourer. Les grosses cellules donnent naissance aux pinacocytes, les petites aux choanocytes et les deux couches de cellules produisent les cellules amiboïdes de la mésoglée. Ceci contraste fortement avec ce qui se produit chez les vrais métazoaires. Chez eux, en effet, ce sont assez typiquement les grosses cellules qui deviennent internes. Voici encore une observation qui différencie les Spongiaires des autres animaux pluricellulaires.
La reproduction asexuée
Certaines éponges marines vivent plusieurs années. Celles des régions tempérées sont généralement dormantes l'hiver et survivent sous une forme réduite dépourvue de choanocytes. Les éponges d'eau douce, qui peuvent faire face à des conditions très difficiles en hiver, survivent rarement au-delà d’une année. Pour perpétuer l'espèce malgré les rigueurs de l'hiver, elles disposent de cellules particulières: les gemmules.
Une gemmule typique est composée d’un amas d'archéocytes bourrés de réserves nutritives, et entourés de cellules amiboïdes. Parmi celles-ci, les unes fabriquent la double coque dure de spongine, d'autres produisent des spicules. Ces protections permettent aux gemmules de résister au gel et à la sécheresse éventuelle. Au printemps, les archéocytes reprennent une vie active, se multiplient, se différencient, sortent de la gemmule et se développent en une nouvelle éponge adulte. Il s'agit ici d'une multiplication asexuée, de nombreuses gemmules étant fabriquées par une même éponge.
Le bourgeonnement
Un autre mode de reproduction asexuée est la formation de bourgeons à partir d’un individu. Selon les cas, ces bourgeons forment des individus qui restent associés à l’individu originel ou qui s’en détachent. S’ils restent associés, le bourgeonnement aboutit à la formation de colonies.

La régénération
Un phénomène qui peut être rapproché de la reproduction asexuée, c'est la régénération. La régénération résulte d'un traumatisme. Ce processus est employé dans la production des éponges naturelles que l’on peut trouver dans le commerce. Une éponge est fragmentée en de nombreux petits morceaux qui sont fixés à une pierre puis immergés. La régénération et quelques années de croissance finissent par produire des éponges de taille commercialisable.
Des expériences maintenant classiques démontrent cette capacité étonnante de régénération. Si on force les tissus vivants d'une éponge à passer au travers d'un tamis à très fines mailles, on sépare ainsi les différentes cellules. Les cellules séparées se réorganisent après une courte période et reforment elles-mêmes de nouvelles éponges. Quelques facteurs indispensables à cette réorganisation ont été mis en évidence. Il s'agit des ions calcium et magnésium, participant aux interactions des molécules permettant l'adhésion entre cellules et aux protéines de la matrice extracellulaire. Ce modèle est utilisé en biologie fondamentale pour étudier des phénomènes d'interactions cellulaires.
Exercices :
Pour rassembler vos idées, mettez par écrit les caractéristiques principales qui vous paraissent bien définir les Spongiaires.
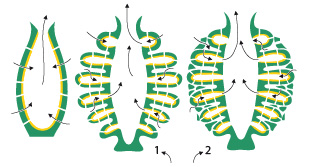
3. Diversité du groupe, origine et évolution - 3.1. Diversité du groupe
Vous voyez ici les variations de structure chez les Eponges : en jaune, le revêtement interne de choanocytes formant des canaux dans l’animal et permettant ainsi les mouvements d’eau ; en vert, le revêtement externe et la mésoglée. De gauche à droite, on dénomme ces types de structure d’éponges ascon, sycon et leucon.
Une coupe au travers d’une éponge permet de révéler leur structure creusée de canaux. Chez les formes les plus primitives, de type ascon, la cavité centrale aux parois simples et percées de petits orifices communique avec l’extérieur par un large orifice, l’oscule, . L’eau provenant de l’extérieur entre par ces petits orifices, les pores inhalants, puis est évacuée par l’oscule. Cette structure ascon impose une forte limitation de la taille de l'animal. En effet, le courant d'eau est ralenti dans la cavité centrale et peut s’arrêter si celle-ci contient trop d'eau pour qu'elle puisse être déplacée rapidement vers l'oscule par les cellules flagellées, les choanocytes, qui bordent la face interne de la cavité. L'augmentation du volume de la cavité centrale ne serait pas accompagnée d'un accroissement suffisant de la surface des choanocytes pour rectifier le problème. Ainsi, les éponges de type ascon sont-elles toujours petites, d’une taille nettement inférieure au centimètre.
Au cours de l’évolution des éponges, la paroi du corps s'est plissée, et la cavité centrale s'est réduite au cours du développement. Les éponges qui montrent les premiers stades de plissement sont appelées sycon. Chez elles, la paroi du corps est plissée horizontalement et a formé des évaginations en doigts de gant. Dans les éponges de ce type, les choanocytes sont confinés dans les évaginations appelées canaux à choanocytes. Les invaginations qui donnent vers l’extérieur sont dénommées canaux inhalants, et sont couvertes d'un tégument de pinacocytes. Les canaux inhalants s'ouvrent dans les canaux à choanocytes par des ouvertures équivalentes aux pores inhalants des éponges ascon. Les flèches sur le schéma montrent le trajet de l'eau. Le plissement accroît la surface des tapis des choanocytes et la réduction de la cavité centrale diminue le volume d'eau qui doit être mis en circulation. Une plus grande taille de l’éponge devient alors possible.
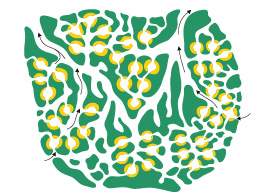
Leucon
Un stade encore plus spécialisé de la structure leucon se développe par le remplissage de l'extrémité ouverte des canaux inhalants avec de la mésoglée et une couverture de pinacocytes. Des ouvertures persistent pour l'entrée de l'eau. Ce type est appelé leucon. Chez les éponges de type leucon, le plus haut degré de complexité de la paroi est atteint. Les canaux de choanocytes se sont eux-mêmes plissés ou évaginés pour former de petites chambres sphériques. Dans cette structure, la cavité centrale a généralement disparu, et seuls subsistent des canaux qui mènent à un oscule unique ou à plusieurs. Le nombre de chambres à choanocytes peut être énorme. Microciona par exemple, contient dix mille chambres par millimètre cube, chacune ayant un diamètre de 30 micromètres environ et contenant une cinquantaine de choanocytes. La plupart des éponges sont construites sur le plan leucon, ce qui montre l'efficacité de ce type de structure. Ces éponges peuvent atteindre des tailles considérables et des formes très variées.

3.2. Origine et évolution du groupe
L'origine des éponges pose quelques problèmes très intéressants. L'absence d'organes, le degré peu élevé d'interdépendance des cellules et une différenciation cellulaire peu poussée sont certainement des caractéristiques primitives. Il est donc pertinent de voir là un premier essai de passage à l'état pluricellulaire.
D'autre part, l'architecture du corps construite autour d'un système de canaux est unique dans le monde animal. Les cellules différenciées des éponges sont aussi très particulières, on n'en retrouvera à peu près aucune pareille chez les autres animaux. Ces considérations pourraient suggérer que les éponges dérivent d'un ancêtre différent de celui des autres animaux.

3.3. Une illustration de la démarche scientifique
Dans cette section, nous allons mettre en pratique les principes d’analyses de données scientifiques qui vous ont été proposés dans le texte d’introduction . Différentes informations de type morphologique et moléculaire vous sont d’abord fournies. Ensuite, nous les synthétisons pour vous montrer comment les chercheurs sont parvenus à proposer un scénario plausible, et testable, pour les trois questions qui nous préoccupent ici.
Ces principes d’analyse et de synthèse des données vous seront nécessaires pour réaliser les travaux de groupes en cours d’année, puisqu’ils sont applicables quelle que soit la question de recherche qui vous sera posée. Veillez donc à bien comprendre comment nous procédons dans les dias suivantes.
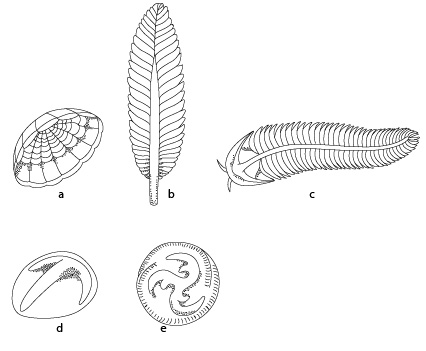
Faune d'Ediacara
Représentation de la faune d’Ediacara. Faune composée de représentants primitifs de groupes plus jeunes, cnidaires (coraux mous et médusoïdes (a- méduse, b-coraux mous), annélides (vers segmentés) ou arthropodes (c- arthropodes nus ou annélides); par contre, (d et e) ne ressemblent à aucun animal connu: Vendobiotes. Faune encore très énigmatique. Dessins extraits de Cloud, P., 1983, La Biosphère. Pour la Science, 73.
L’étude des couches géologiques anciennes à la surface de la terre nous révèle que la première faune complexe pour laquelle on a de bonnes données paléontologiques est la faune d’Ediacara, présente dans des roches qui ont entre 630–540 millions d’années, période qu’on appelle le Précambrien. On y retrouve des traces de fossiles présentant une structure morphologique complètement inédite par rapport à la faune actuelle, mais aussi des traces de fossiles apparentés aux Spongiaires et aux Cnidaires. Cette faune nous révèle que les Spongiaires étaient donc déjà présents longtemps avant le début de la période cambrienne (540 à 490 millions d’années). Il reste cependant surprenant qu’on ne retrouve pas de spicules, typiques des Spongiaires, au Précambrien.
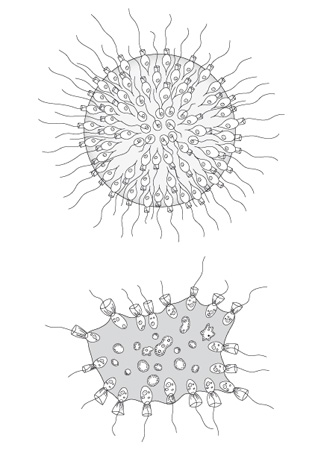
Espèces de Protistes Choanoflagellés
Deux espèces de Protistes Choanoflagellés vivant en groupe : en haut Proterospongia ; en bas Sphaeroeca. Leur organisation ressemble à celle des Spongiaires
Quand on s’intéresse aux spongiaires vivant à l’heure actuelle, on remarque une similarité structurelle remarquable entre les cellules nourricières des Spongiaires, les choanocytes, et un groupe de protistes appelés Choanoflagellés. Les deux types de cellules possèdent un flagelle unique entouré par un collier de fins tentacules (la collerette). On a donc émis l’hypothèse que les Eponges auraient évolué à partir d’un ancêtre de type protiste Choanoflagellé.
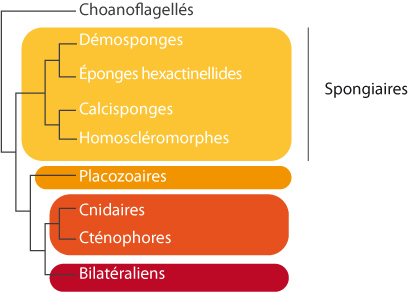
Arbre phylogénétique
Arbre phylogénétique obtenus sur base du séquençage de 129 gènes correspondant à un total de 30257 acides aminés (d’après Telford 2009). Cet arbre montre qu’on distingue 4 groupes d’éponges, et les relations entre ces différents groupes.
Etant donné le faible nombre de caractères morphologiques présents chez les Spongiaires, d’autres chercheurs se sont tournés vers des méthodes de biologie moléculaire, dont le séquençage d’un grand nombre de gènes. Cette méthode a pu se développer depuis le milieu des années 1980 grâce à une avancée technique majeure, l’invention de la PCR (polymerase chain reaction). Le séquençage de différents gènes conservés au sein des Métazoaires et parmi les Spongiaires a montré que ce groupe est formé de quatre groupes distincts :
- les Calcisponges (Calcarea) qui partagent un squelette formé de spicules calcaires, tels que Grantia sp ;
- les Hexactinellides tels que Euplecetella sp qui produisent des spicules de silice à 6 pointes formant une construction complexe mais légère, et sont formés d’un choanosyncitium plutôt que d’un choanoderme, et qui ne présentent pas de pinacoderme continu ;
- les Démosponges comme Halichondria et Haliclona qui possèdent toutes une structure de type leucon et des spicules constitués de silice organisée autour d’une matrice de spongine; environ 90% des éponges font partie de ce groupe ;
- les Homoscléromorphes, le groupe le plus récemment identifié, représentés par une seule famille, les Plakinidae.
Ces différents groupes sont monophylétiques et seront vus en travaux pratiques.
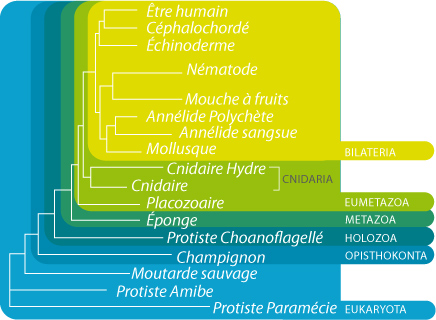
Arbre phylogénétique
Par ailleurs, le premier génome complet d’un Porifère a été séquencé en 2010, c’est celui d'Amphimedon queenslandica (Srivastava et al., 2010 dans Nature). Sur base de l’arbre phylogénétique qui en découle, on constate que les Porifères (Eponge) sont à la base des Métazoaires, et que l’organisme le plus proche des Eponges est un bien un Protiste choanoflagellé, Monosiga sp. Le séquençage du génome complet d'A. queenslandica a aussi permis de montrer que cette éponge possède 18,000 gènes, alors que ce nombre de gènes codant pour des protéines avoisine 25,000 chez l’homme! De façon étonnante peut-être, cette éponge comprend la plupart des gènes utilisés dans la formation des tissus épithéliaux (adhérence cellulaire, communication intercellulaire, protéines kinases,...). Les tissus épithéliaux sont un caractère clef de l’évolution des Métazoaires, étant donné que ce type de tissus est un élément essentiel pour l’élaboration de plans de structures morphologiques complexes et le développement d’organismes de grande taille. Les tissus épithéliaux permettent en effet de créer une compartimentalisation entre organes et entre le corps et le milieu extérieur. Autrefois, on devait se contenter d’observer les caractéristiques des epithelia au microscope. On peut maintenant tester l’existence d’epithelia dans un organisme en analysant la présence et l’expression des gènes impliqués dans la formation de ces epithelia, par exemple les gènes établissant la polarité cellulaire, formant les jonctions intercellulaires, ou établissant et régulant l’adhésion à la lame basale. En effet, ces gènes sont conservés (i.e. sont les mêmes) pour les insectes, les nématodes et les vertébrés, et sont donc vraisemblablement les mêmes pour tous les animaux.
Comme nous l’avons vu, les éponges ne possèdent pas de tissu épithélial à proprement parler et pourtant, possèdent de nombreux gènes qui les produisent chez d’autres groupes taxonomiques. Par contre, on ne retrouve pas la plupart de ces gènes dans les génomes de Protistes. Ces gènes caractéristiques des tissus épithéliaux sont donc apparus au sein de l’ancêtre des Métazoaires, les éponges.
Dans le génome de cette éponge, on a aussi trouvé une série de gènes utilisés par les organismes plus “évolués” pour produire les tissus musculaires et nerveux. Une telle complexité moléculaire est surprenante chez les éponges qui ne possèdent ni muscles ni système nerveux.
Tout ceci indique que l’ancêtre des Eponges était sans doute plus complexe que ce qu’on pensait jusqu’à présent étant donné que ces gènes avaient une fonction (similaire ?) chez cet ancêtre.
Exercice :
On peut maintenant tenter de répondre aux trois questions qui nous préoccupent. Synthétisez vous-même les conclusions qu’on peut tirer pour chacune des trois questions. C’est un bon exercice pour préparer votre travail en groupe.